
Fire Haired14
Banned
- Messages
- 2,185
- Reaction score
- 583
- Points
- 0
- Y-DNA haplogroup
- R1b DF27*
- mtDNA haplogroup
- U5b2a2b1
Fire Haired(me): I am sincerely sorry and embarrassed for how arrogant and excessive I was at this forum. I have matured since then, and I'm ready to become a member again. I am requesting to regain my membership because I want to learn through conversation and from people who are educated and great critical thinkers-workers-researchers. Trust me i won't make threads like R1b L51-L11 Germanic Italo Celts: Rulers and conqueres of Bronze-Iron age west Europe again. I believe that at Eupedia I will improve much faster.
Ancient individuals:
Hunter gatherers: Loschbour(Loschbour, Luxembourg 8000BP), La Brana-1(7940-7690 cal BP, northern Spain), Motala2-3-6-9-12(Motala, Sweden 8000BP), Skoglund_HG(Gotland, Sweden 4,000-4,800BP), MA1(Mal'ta, Siberia 24,000BP), AG2(near MA1, 17,000BP).
Farmers: Otzi(Otztal, Alps 5,225BP), Stuttgart(Stuttgart, Germany 7,000BP), Skohlund_farmer(Gokhem, Sweden 5,000BP).
Click here to see a google map I made of the Mesolithic DNA from at this site.
Relationships of the ancient genomes to each other
Supplementary Information 14 begins with testing how related the ancient genomes(Loschbour, La Brana-1, Motala12, Stuttgart, MA1) are to each other using f4-statistics.
I have a weak understanding of all the science behind this, but anyone can understand how to interpret the results after reading this paragraph thoroughly.
Here are some obvious findings from the tests. All of the European hunter gatherers are closer to each other than to Stuttgart and MA1, Loschbour and La Brana-1 are closest to each other, MA1 and Stuttgart are closer to the European hunter gatherers than to each other, Stuttgart is closer to Loschbour and Motala12 than to La Brana-1, Motala12 has ANE(MA1 related) ancestry, and all of the European hunter gatherers are a little bit closer to Stuttgart than to MA1 except Motala12 who is slightly closer to MA1 than to Stuttgart.
The authors make a big deal about Loschbour and La Brana-1's results being constant with them descending from the same west European hunter gatherer population but they never mentioned the obvious relation between Loschbour and Motala12. The statistic f4(Loschbour, Chimp, La Brana-1, Motala12) is much less significantly positive (5.962) than f4(La Brana-1, Chimp, Loschbour, Motala12) which scores 11.324. So La Brana-1 is much more distant from Motala12 than Loschbour is. La Brana-1 and Loschbour score very similar results unless Motala12 is involved, in which Loschbour always shows a much closer relation to Motala12 than La Brana-1 does.
If you took out Motala12's ANE ancestry he would probably be almost as close to Loschbour as La Brana-1 is. In my opinion Loschbour had almost as much central European specific hunter gatherer ancestry as he did La Brana-1 related west European hunter gatherer ancestry. Even though Motala12 lived in Sweden many of his ancestors lived in central Europe probably around where Loschbour lived. The fact that all of the Swedish hunter gatherers sampled in this study belonged(and could successfully find their Y DNA haplogroup) to Y DNA I and that Motala12 and Motala3 had many of the same Y DNA I2a1b mutations as Loschbour proves they have common paternal ancestors who probably lived in central Europe.
It is also important to note that Neolithic hunter gatherers from Gotland(Skoglund_HG) cluster very closely to the Mesolithic Swedish hunter gatherer(Motala, not very far from Gotland) in the PCAs, which means there was strong genetic continuum in Scandinavian hunter gatherers from the Mesolithic-Neolithic.
Stuttgart is closer to both Motala12(despite his ANE ancestry) and Loschbour than she is to La Brana-1, even though La Brana-1 had some farmer ancestry. This is probably because LBK farmers mixed with European hunter gatherers from the Balkans and central Europe who were more related to Loschbour and Motala12 than to Iberian(south-western edge of Europe) hunter gatherers like La Brana-1.
I am very surprised that all of the European hunter gatherers except Motala12(who had ANE ancestry) are more related to Stuttgart than to MA1. This is because La Brana-1 is more related to MA1 than any modern populations are, west Asian components(mixture of near eastern and ANE) in admixtures are closest to mainly Mesolithic descended north European components, and how closely related La Brana-1 and MA1 seem to be in Davidski aka Polako's PCA's and admixtures(click here and here).
Loschbour and La Brana-1 are not very significantly closer to Stuttgart than to MA1, and Loschbour is a bit closer than La Brana-1 is because Stuttgart has hunter gatherer ancestors who were more related to Loschbour than to La Brana-1. Loschbour and La Brana-1 are probably more related to MA1 than to Stuttgart's near eastern ancestors, but her non basal Eurasian near eastern ancestors were probably more related to Loschbour and La Brana-1 than MA1 is. Stuttgart's near eastern ancestors(west Eurasian and basal Eurasian) are the reason why MA1 is less related to her than to European hunter gatherers and there is a low chance MA1 is more related to her non basal Eurasian near eastern ancestors than to European hunter gatherers. This probably means there was admixture at some point between Caucasoid aka west Eurasian(maybe not accurate definitions) near easterns, Europeans, and east Europeans-Siberians before the Neolithic.
The TreesAfter comparing the ancient genomes to each other and non west Eurasian populations using f4-statistics the authors attempted to make tree models that are consistent with the f-statistics using Loschbour, Stuttgart, MA1, Karitiana, and Onge. I am pretty sure that last sentence is accurate but even if it is not that doesn't change the message of their trees.
After many attempts they finally succeeded with the tree in Figure S14.6., and then used that to place other ancient samples and modern Europeans and near easterns in the tree. La Brana-1 fits as a brotherclade to Loschbour and Otzi fits as a brotherclade with Stuttgart(Figure S14.7). Motala12 fits as an admixture of Loschbour(81%) and MA1(19%)(Figure S14.8).
Stuttgart is fit as an admixture of basal Eurasian(ranges from 32-39% in the trees) and a brothercalde to Loschbour named Y(ranges from 61-66% in the trees). Which means the majority of her ancestry was from west asians who were closely related to Mesolithic Europeans combined with Mesolithic European ancestry.
When modern Europeans are forced to be a 2-way admixture of Loschbour and Stuttgart only Basques, Spanish_North, and Sardinians could successfully fit. There was a picture of a tree with Sardinians shown as as mixture of Loschbour(21%) and Stuttgart(79%). Sardinians are the closest modern relatives to early European farmers but they do have more hunter gatherer ancestry and in one of Davidski's admixtures they score on average 33.3%(click here), which makes sense since Stuttgart had some European hunter gatherer ancestry.
Since most modern Europeans show evidence of having a mixture of near eastern, Mesolithic European, and ANE(MA1-related) ancestry in their PCAs and f-statistics the authors attempted to fit modern Europeans as a mixture of Motala12(who had some ANE ancestry) and Stuttgart. Only Basque, French_South, and Sardinian could successfully fit. Meaning European hunter gatherers with the same amount of ANE ancestry as Motala12 are not enough to explain how high ANE is in most modern Europeans. There was also a picture of Sardinians being fit as an admixture of Motala12(12%) and Stuttgart(88%). They scored much less hunter and much more farmer than in the previous tree probably because they have little to no ANE ancestry.
When they attempted to fit modern Europeans as a mix of Loschbour, MA1, and Stuttgart, 26 European populations fit(not sure how many were tested) and no near eastern populations fit(and definitely no one but Europeans).
They listed many European population's EEF(Stuttgart), WHG(Loschbour), and ANE(MA1) results. The distribution of EEF, WHG, and ANE are 100% consistent with what has already been theorized by many on the distribution of Mesolithic European and near eastern ancestry in modern Europe, but ANE is a third element that was not considered until MA1's genome was sampled.
The authors also used modern near eastern populations without substantial African ancestry(<1%) in the admixture at K=6 and who score their most significant f3-statistic with the pairing of Stuttgart, MA1 in the same model but as a mixture of near eastern(NE) and MA1(ANE). Only five near eastern populations fit the criteria to be put in this new tree: Abkhasian, Chechen, Cypriot, Druze, and Lezgin. Click here to see estimated ANE levals in asia.
Here is the tree they used to estimate NE and ANE ancestry in modern near easterns. I copied and pasted it from the blog For what they were... we are, since i can't copy and paste straight from Laz.
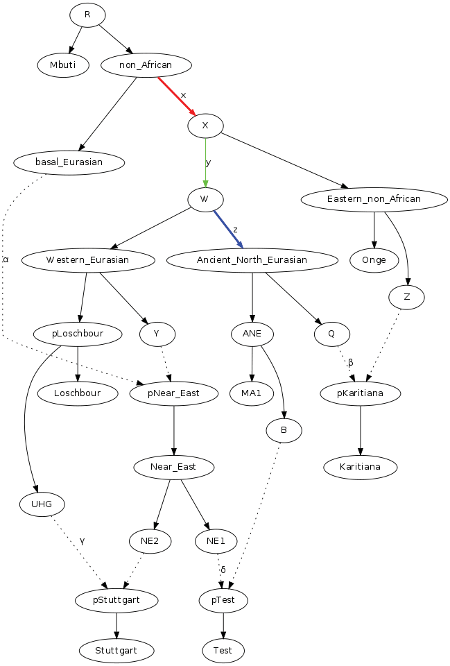
Near eastern is shown as a brotherclade to Loschbour, meaning non basal Eurasian west Asians were very closely related to Mesolithic Europeans.
Here is what the authors have to say about it.
An interesting detail of Fig. S14.20 is that the Near_East is modeled as a mixture of basal_Eurasian and a node Y which forms a clade with Loschbour. Present-day Near Eastern populations are indeed more closely related to European hunter-gatherers than to MA1 despite having some MA1-related ancestry. This can be easily seen in Extended Data Fig. 6C where the range of the statistic f4(Test, Chimp; MA1, Loschbour) is negative for all West Eurasian populations including all Near Eastern ones, suggesting that they share more drift with Loschbour than with MA1 (the statistic is Z<-4 for all West Eurasian populations except the Lezgin where it is Z=-3.6). If we attempt to fit Near Eastern
Here are the NE and ANE results for Abkhasian, Chechen, Cypriot, Druze, and Lezgin
ANE ancestry is higher in much of the near east than in Europe but MA1 is still shares more drift with many Europeans because near easterns have a higher amount of basal Eurasian ancestry.
European's Mesolithic aka WHG ancestry which lacks any basal Eurasian ancestry is the reason why they share more drift with MA1 than near easterns who have more ANE(MA1 related) ancestry.
Possibly this can explain why it seems based on Davidski's work that Mesolithic Europeans and MA1 are brotherclades. If you took near easterns basal Eurasian ancestry out of them they would be a brotherclade to Mesolithic Europeans and MA1 would be their cousin.
Y DNA


Loschbour, 6,000BC Loschbour Luxembourg: Y DNA=pre-I2a1b or brother lineage to I2a1b(I L41+, I PF3742+, I M258+, I M170+, I P389+, I2 L68+, I2 M438+, I2a L460+, I2a1 P37.2+, I2a1b M423+, I2a1b CTS8239+, I2a1b CTS7218+, I2a1b CTS54985+, I2a1b L178+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b CTS5375-, I2a1b CTS8486-, I2a1b1 M359.2-, I2a1b2 L161.1, I2a1b3 L621-)
Motala2, 6,000BC Motala Sweden: Y DNA=I* (I P38+, I PF3742+, I L41+, I1 S108-, I1 L845-, I1 M253-, I2a1b CT1293-, I2a2 L37-)
Motala3 6,000BC Motala Sweden: Y DNA=I2a1b*(I M258+, I PF3742+, I2 L68+, I2a1 P37.2+, I2a1b CTS7218+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b1 M359.2-, I2a1b3 L621-)
Motala6 6,000BC Motala Sweden: Y DNA=? (Q1 L232- Q1a2a L55+)
Motala9 6,000BC Motala Sweden: Y DNA=I* (I P38+, I1 P40-)
Motala12 6,000BC Motala Sweden: Y DNA=pre-I2a1b or brother lineage to I2a1b(I PF3742+, I M258+, I M170+, I2 L68+, I2a L460+, I2a1 P37.2+, I2a1b CTS7218+, I2a1b CTS5985+. I2a1b L178+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b CTS5375-, I2a1b CTS8486-, I2a1b1 M359.2-, I2a1b3 L621-)
New Y chromosome SNP's were tested for Loschbour, Motala2, Motala3, and Motala12. Loschbour and Motala-2-3-9-12 were all tested and found to have at least one defining mutation of Y DNA I. Motala6 is Q1a2a L55+ but Q1 L232-, so his Y DNA haplogroup is unknown. Loschbour was tested for 9 mutations that define I2a1b and had 7, Motala12 was tested for 7 and had 5, and Motala3 was tested for 3 and had all 3. Motala12 and Loschbour were missing the same two I2a1b defining mutations(CTS5375, CTS8486). Possibly Motala12 and Loschbour belonged to a brother lineage or an ancestral version of I2a1b.
Motala6 did not have any I1 defining mutations that he was tested for and he did not have I2a1b alleles in SNP CTS1293(only I2a1b defining SNP he was tested for) unlike Motala12, Loschbour, and Motala3, so all that is known about Motala6's Y DNA is that he was a member of haplogroup I. Motala9 was tested for an I1 mutation and was negative but he was not tested for any I2(including all subclades) mutations.
There are many Y DNA I2a1 subclades in Europe today, it's most diverse in western Europe and all are probably native(Mesolithic) to western Europe. I2a1b1 L621 is very popular in south-eastern Europe but i think is descended of post Mesolithic west Europeans. I2a2 and I1 all probably descend from central-west Mesolithic Europeans. I1(which has 25 defining mutations) may not have existed in the Mesolithic but it's ancestral version certainly did, probably somewhere in central Europe. I2a2 was largely dispersed by Celts and Germans, but it did exist somewhere in Mesolithic central-west Europe. For more about Y DNA I today: Eupedia Y DNA I1, Eupedia Y DNA I2
New SNP's associated with pigmentation were tested for Loschbour, Stuttgart, and Motala12(Motala12 was not tested at all in their previous preprint). The same 8plex and Hirisplex were listed for Loschbour and Stuttgart, but some results were also listed for Motala12.
Here are the predictions for hair and eye color of Loschbour and Stuttgart using the Hirisplex model.
Motala12(Sweden, 6,000BC) like Loschbour(Luxembourg, 6220-5990 BC) and La brana-1(Spain, 5940-5690BC) most likely had light(prob. blue) eyes and dark hair. That's 3/3 for Mesolithic Europeans which means that combination was widespread and very popular in Europe by at least 8,000 years ago. The most compelling evidence Motala12 also had light eyes(probably blue) is that he had G/G alleles in SNP rs12913832.
Motala12 had T/T alleles in SNP rs12203592(in gene IRF4) like Loschbour and La Brana-1(T/C). Today T/T and T/C alleles in that SNP are very rare in modern Europeans and absent in east asians and Africans(see here). IRF4 is associated with sensitivity of skin to sun exposure, freckles, blue eyes, and brown hair color(got that from here).
Alot of the stuff about pigmentation is new to me so I am going to give some quotes.
Motala12 was tested for 10 of 13 H-1 HERC2/OCA2 haplotype SNPs and had all of them except in rs3935591 he had T/C. 97% of modern blue eyed people(and Loschbour) have C/C. Of the mutations tested and associated with blonde and red hair none of the ancient samples had any.
Motala12 surprisingly had alleles A/A('light skin" version) in SNP rs1426654(in gene SLC24A5) like near eastern(main ancestors migrated to Europe) farmers Stuttgart and Otzi and unlike fellow European hunter gatherers Loschbour and La Brana-1 who had G/G.
There is another similarity between Motala12, near eastern farmers, and most modern west Eurasians with SNP's associated with skin color.
This means that middle eastern ancestors are not the sole source of the derived rs1426654 pigmentation-lightening allele in modern Europeans. My opinion is that the skin color of these ancient hunter gatherers is unknown. A good guess is that they had dark skin, but there is evidence in modern people that they had light skin and possibly some(in eastern Europe?) who have not been sampled had blonde and red hair. The farmers were probably as light as Sardinians(their closest relatives), as dark as west Asians, or somewhere in-between. It is pretty certain that both European hunter gatherers and near eastern farmers had primarily dark hair, but the hunter gatherers were mainly light eyed while the farmers were mainly dark eyed.
Of the three mutations(in genes TYR, SLC24A5, and SLC45A2) most associated with light skin in Europe(all are just as popular in west Asians except the one in gene SLC45A2 which is ~50% in west Asians and ~100% in Europeans) La brana-1 had 0/3, Loschbour had 0/3, Motala12 had 1/3, Stuttgart had 2/3, and of tested Otzi had 2/2. Copper age(Yamna and Catacomb cultures) and Eneolithic people of the Pontiac steppe have been found to most likely have had around 90% brown eyes, an even higher percentage of dark hair, and possibly dark skin(click here). In contrast bronze and iron age Indo Iranians of south Siberia who are their supposed descendants and relatives had majority light eyes, light hair, and probably light skin(of SNP's associated with pigmentation tested their frequencies were no different from modern northern and eastern Europeans).
The distribution of light eyes and dark eyes in modern Europeans is nearly identical to the distribution of Mesolithic and near eastern ancestry, which makes sense since so far 3 out of 3 Mesolithic Europeans tested have light eyes and 2 out of 2 early European farmers tested so far have dark eyes. I think light eyes in modern European's ancestors probably went through powerful selection after the farmers and hunter gatherers mixed but the mutations for it mainly came from their hunter gatherer ancestors.
So far 3 out of 3 Mesolithic Europeans and 2 out of 2 Neolithic Europeans tested fo SNPs that can accurately determine hair color have no mutations associated with blonde or red hair. It would make sense that high percentages of light hair originated in Mesolithic Europeans since they were mainly light eyed(light haired people have a much higher percentage of light eyes). I think it is likely that light hair did exist in Mesolithic Europeans(and early farmers) just because they were west Eurasian but that doesn't mean a high percentage of them did. The origin of a high percentage of non dark hair in modern Europeans(unlike any other people in the world) is still a mystery.
Ancient individuals:
Hunter gatherers: Loschbour(Loschbour, Luxembourg 8000BP), La Brana-1(7940-7690 cal BP, northern Spain), Motala2-3-6-9-12(Motala, Sweden 8000BP), Skoglund_HG(Gotland, Sweden 4,000-4,800BP), MA1(Mal'ta, Siberia 24,000BP), AG2(near MA1, 17,000BP).
Farmers: Otzi(Otztal, Alps 5,225BP), Stuttgart(Stuttgart, Germany 7,000BP), Skohlund_farmer(Gokhem, Sweden 5,000BP).
Click here to see a google map I made of the Mesolithic DNA from at this site.
Relationships of the ancient genomes to each other
Supplementary Information 14 begins with testing how related the ancient genomes(Loschbour, La Brana-1, Motala12, Stuttgart, MA1) are to each other using f4-statistics.
I have a weak understanding of all the science behind this, but anyone can understand how to interpret the results after reading this paragraph thoroughly.
We first report (Table S14.1) statistics of the form f4(Ancient1, Chimp; Ancient2, Ancient3) for the ancient samples: Loschbour, Stuttgart, Motala12, MA1, and LaBrana. Such statistics determine whether (Ancient2, Ancient3) are consistent with being a clade relative to Ancient1. If they are not a clade, the statistic shows whether Ancient1 is more closely related to Ancient2 (in which case it is positive), or Ancient3 (in which case it is negative).
Here are some obvious findings from the tests. All of the European hunter gatherers are closer to each other than to Stuttgart and MA1, Loschbour and La Brana-1 are closest to each other, MA1 and Stuttgart are closer to the European hunter gatherers than to each other, Stuttgart is closer to Loschbour and Motala12 than to La Brana-1, Motala12 has ANE(MA1 related) ancestry, and all of the European hunter gatherers are a little bit closer to Stuttgart than to MA1 except Motala12 who is slightly closer to MA1 than to Stuttgart.
The authors make a big deal about Loschbour and La Brana-1's results being constant with them descending from the same west European hunter gatherer population but they never mentioned the obvious relation between Loschbour and Motala12. The statistic f4(Loschbour, Chimp, La Brana-1, Motala12) is much less significantly positive (5.962) than f4(La Brana-1, Chimp, Loschbour, Motala12) which scores 11.324. So La Brana-1 is much more distant from Motala12 than Loschbour is. La Brana-1 and Loschbour score very similar results unless Motala12 is involved, in which Loschbour always shows a much closer relation to Motala12 than La Brana-1 does.
If you took out Motala12's ANE ancestry he would probably be almost as close to Loschbour as La Brana-1 is. In my opinion Loschbour had almost as much central European specific hunter gatherer ancestry as he did La Brana-1 related west European hunter gatherer ancestry. Even though Motala12 lived in Sweden many of his ancestors lived in central Europe probably around where Loschbour lived. The fact that all of the Swedish hunter gatherers sampled in this study belonged(and could successfully find their Y DNA haplogroup) to Y DNA I and that Motala12 and Motala3 had many of the same Y DNA I2a1b mutations as Loschbour proves they have common paternal ancestors who probably lived in central Europe.
It is also important to note that Neolithic hunter gatherers from Gotland(Skoglund_HG) cluster very closely to the Mesolithic Swedish hunter gatherer(Motala, not very far from Gotland) in the PCAs, which means there was strong genetic continuum in Scandinavian hunter gatherers from the Mesolithic-Neolithic.
Stuttgart is closer to both Motala12(despite his ANE ancestry) and Loschbour than she is to La Brana-1, even though La Brana-1 had some farmer ancestry. This is probably because LBK farmers mixed with European hunter gatherers from the Balkans and central Europe who were more related to Loschbour and Motala12 than to Iberian(south-western edge of Europe) hunter gatherers like La Brana-1.
I am very surprised that all of the European hunter gatherers except Motala12(who had ANE ancestry) are more related to Stuttgart than to MA1. This is because La Brana-1 is more related to MA1 than any modern populations are, west Asian components(mixture of near eastern and ANE) in admixtures are closest to mainly Mesolithic descended north European components, and how closely related La Brana-1 and MA1 seem to be in Davidski aka Polako's PCA's and admixtures(click here and here).
Loschbour and La Brana-1 are not very significantly closer to Stuttgart than to MA1, and Loschbour is a bit closer than La Brana-1 is because Stuttgart has hunter gatherer ancestors who were more related to Loschbour than to La Brana-1. Loschbour and La Brana-1 are probably more related to MA1 than to Stuttgart's near eastern ancestors, but her non basal Eurasian near eastern ancestors were probably more related to Loschbour and La Brana-1 than MA1 is. Stuttgart's near eastern ancestors(west Eurasian and basal Eurasian) are the reason why MA1 is less related to her than to European hunter gatherers and there is a low chance MA1 is more related to her non basal Eurasian near eastern ancestors than to European hunter gatherers. This probably means there was admixture at some point between Caucasoid aka west Eurasian(maybe not accurate definitions) near easterns, Europeans, and east Europeans-Siberians before the Neolithic.
The Trees
After many attempts they finally succeeded with the tree in Figure S14.6., and then used that to place other ancient samples and modern Europeans and near easterns in the tree. La Brana-1 fits as a brotherclade to Loschbour and Otzi fits as a brotherclade with Stuttgart(Figure S14.7). Motala12 fits as an admixture of Loschbour(81%) and MA1(19%)(Figure S14.8).
Stuttgart is fit as an admixture of basal Eurasian(ranges from 32-39% in the trees) and a brothercalde to Loschbour named Y(ranges from 61-66% in the trees). Which means the majority of her ancestry was from west asians who were closely related to Mesolithic Europeans combined with Mesolithic European ancestry.
When modern Europeans are forced to be a 2-way admixture of Loschbour and Stuttgart only Basques, Spanish_North, and Sardinians could successfully fit. There was a picture of a tree with Sardinians shown as as mixture of Loschbour(21%) and Stuttgart(79%). Sardinians are the closest modern relatives to early European farmers but they do have more hunter gatherer ancestry and in one of Davidski's admixtures they score on average 33.3%(click here), which makes sense since Stuttgart had some European hunter gatherer ancestry.
Since most modern Europeans show evidence of having a mixture of near eastern, Mesolithic European, and ANE(MA1-related) ancestry in their PCAs and f-statistics the authors attempted to fit modern Europeans as a mixture of Motala12(who had some ANE ancestry) and Stuttgart. Only Basque, French_South, and Sardinian could successfully fit. Meaning European hunter gatherers with the same amount of ANE ancestry as Motala12 are not enough to explain how high ANE is in most modern Europeans. There was also a picture of Sardinians being fit as an admixture of Motala12(12%) and Stuttgart(88%). They scored much less hunter and much more farmer than in the previous tree probably because they have little to no ANE ancestry.
When they attempted to fit modern Europeans as a mix of Loschbour, MA1, and Stuttgart, 26 European populations fit(not sure how many were tested) and no near eastern populations fit(and definitely no one but Europeans).
They listed many European population's EEF(Stuttgart), WHG(Loschbour), and ANE(MA1) results. The distribution of EEF, WHG, and ANE are 100% consistent with what has already been theorized by many on the distribution of Mesolithic European and near eastern ancestry in modern Europe, but ANE is a third element that was not considered until MA1's genome was sampled.
| EEF | WHG | ANE | |
| Albanian | 78.1 | 9.2 | 12.7 |
| Ashkenazi_Jew | 93.1 | 0 | 6.9 |
| Basque | 59.3 | 29.3 | 11.4 |
| Belarusian | 41.8 | 43.1 | 15.1 |
| Bergamo | 71.5 | 17.7 | 10.8 |
| Bulgarian | 71.2 | 14.7 | 14.1 |
| Croatian | 56.1 | 29.3 | 14.5 |
| Czech | 49.5 | 33.8 | 16.7 |
| English | 49.5 | 36.4 | 14.1 |
| Estonian | 32.2 | 49.5 | 18.3 |
| French | 55.4 | 31.1 | 13.5 |
| French_South | 67.5 | 19.5 | 13 |
| Greek | 79.2 | 5.8 | 15.1 |
| Hungarian | 55.8 | 26.4 | 17.9 |
| Icelandic | 39.4 | 45.6 | 15 |
| Lithuanian | 36.4 | 46.4 | 17.2 |
| Maltese | 93.2 | 0 | 6.8 |
| Norwegian | 41.1 | 42.8 | 16.1 |
| Orcadian | 45.7 | 38.5 | 15.8 |
| Sardinian | 81.7 | 17.5 | 0.8 |
| Scottish | 39 | 42.8 | 18.2 |
| Sicilian | 90.3 | 0 | 9.7 |
| Spainish | 80.9 | 6.8 | 12.3 |
| Spainish_North | 71.3 | 12.5 | 16 |
| Tuscan | 74.6 | 13.6 | 11.8 |
| Ukrainian | 46.2 | 38.7 | 15.1 |
The authors also used modern near eastern populations without substantial African ancestry(<1%) in the admixture at K=6 and who score their most significant f3-statistic with the pairing of Stuttgart, MA1 in the same model but as a mixture of near eastern(NE) and MA1(ANE). Only five near eastern populations fit the criteria to be put in this new tree: Abkhasian, Chechen, Cypriot, Druze, and Lezgin. Click here to see estimated ANE levals in asia.
Here is the tree they used to estimate NE and ANE ancestry in modern near easterns. I copied and pasted it from the blog For what they were... we are, since i can't copy and paste straight from Laz.
Near eastern is shown as a brotherclade to Loschbour, meaning non basal Eurasian west Asians were very closely related to Mesolithic Europeans.
Here is what the authors have to say about it.
An interesting detail of Fig. S14.20 is that the Near_East is modeled as a mixture of basal_Eurasian and a node Y which forms a clade with Loschbour. Present-day Near Eastern populations are indeed more closely related to European hunter-gatherers than to MA1 despite having some MA1-related ancestry. This can be easily seen in Extended Data Fig. 6C where the range of the statistic f4(Test, Chimp; MA1, Loschbour) is negative for all West Eurasian populations including all Near Eastern ones, suggesting that they share more drift with Loschbour than with MA1 (the statistic is Z<-4 for all West Eurasian populations except the Lezgin where it is Z=-3.6). If we attempt to fit Near Eastern
Here are the NE and ANE results for Abkhasian, Chechen, Cypriot, Druze, and Lezgin
| Near East | ANE(fitted) | ANE(lower bound) | |
| Abkhasian | 81.4 | 18.6 | 15.7 ± 5.2 |
| Chechen | 73 | 27 | 24.4 ± 4.9 |
| Cyproit | 86.7 | 13.3 | 9.7 ± 5.6 |
| Durze | 88.2 | 11.8 | 4.7 ± 5.5 |
| Lezgin | 71.2 | 28.8 | 26.1 ± 4.9 |
ANE ancestry is higher in much of the near east than in Europe but MA1 is still shares more drift with many Europeans because near easterns have a higher amount of basal Eurasian ancestry.
The high affinity of the Northeast Caucasus to MA1 is also demonstrated in Extended Data Fig. 6, where the statistic f4(Test, Chimp; MA1, Loschbour) exhibits highest values in the region. In light of our other results, it is not surprising that these populations would have high ANE-related ancestry. They are at the northern end of the Near Eastern cline (Fig. 1B) and have the highest values of common genetic drift with MA1 among Near Eastern populations (Extended Data Fig. 4), as measured by f4(Test, Stuttgart; MA1, Chimp). However, the high MA1-related admixture in Northeast Caucasians seemingly contradicts Extended Data Fig. 4 which shows many Europeans to have even higher values of the statistic.
European's Mesolithic aka WHG ancestry which lacks any basal Eurasian ancestry is the reason why they share more drift with MA1 than near easterns who have more ANE(MA1 related) ancestry.
Intuitively, the shared drift shared between a test population and MA1 is diluted by Near Eastern ancestry (because of the Basal Eurasian ancestry in the Near East), and augmented by WHG ancestry (because of the lack of Basal Eurasian ancestry in Loschbour).
Possibly this can explain why it seems based on Davidski's work that Mesolithic Europeans and MA1 are brotherclades. If you took near easterns basal Eurasian ancestry out of them they would be a brotherclade to Mesolithic Europeans and MA1 would be their cousin.
Y DNA
Loschbour, 6,000BC Loschbour Luxembourg: Y DNA=pre-I2a1b or brother lineage to I2a1b(I L41+, I PF3742+, I M258+, I M170+, I P389+, I2 L68+, I2 M438+, I2a L460+, I2a1 P37.2+, I2a1b M423+, I2a1b CTS8239+, I2a1b CTS7218+, I2a1b CTS54985+, I2a1b L178+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b CTS5375-, I2a1b CTS8486-, I2a1b1 M359.2-, I2a1b2 L161.1, I2a1b3 L621-)
Motala2, 6,000BC Motala Sweden: Y DNA=I* (I P38+, I PF3742+, I L41+, I1 S108-, I1 L845-, I1 M253-, I2a1b CT1293-, I2a2 L37-)
Motala3 6,000BC Motala Sweden: Y DNA=I2a1b*(I M258+, I PF3742+, I2 L68+, I2a1 P37.2+, I2a1b CTS7218+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b1 M359.2-, I2a1b3 L621-)
Motala6 6,000BC Motala Sweden: Y DNA=? (Q1 L232- Q1a2a L55+)
Motala9 6,000BC Motala Sweden: Y DNA=I* (I P38+, I1 P40-)
Motala12 6,000BC Motala Sweden: Y DNA=pre-I2a1b or brother lineage to I2a1b(I PF3742+, I M258+, I M170+, I2 L68+, I2a L460+, I2a1 P37.2+, I2a1b CTS7218+, I2a1b CTS5985+. I2a1b L178+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b CTS5375-, I2a1b CTS8486-, I2a1b1 M359.2-, I2a1b3 L621-)
New Y chromosome SNP's were tested for Loschbour, Motala2, Motala3, and Motala12. Loschbour and Motala-2-3-9-12 were all tested and found to have at least one defining mutation of Y DNA I. Motala6 is Q1a2a L55+ but Q1 L232-, so his Y DNA haplogroup is unknown. Loschbour was tested for 9 mutations that define I2a1b and had 7, Motala12 was tested for 7 and had 5, and Motala3 was tested for 3 and had all 3. Motala12 and Loschbour were missing the same two I2a1b defining mutations(CTS5375, CTS8486). Possibly Motala12 and Loschbour belonged to a brother lineage or an ancestral version of I2a1b.
Motala6 did not have any I1 defining mutations that he was tested for and he did not have I2a1b alleles in SNP CTS1293(only I2a1b defining SNP he was tested for) unlike Motala12, Loschbour, and Motala3, so all that is known about Motala6's Y DNA is that he was a member of haplogroup I. Motala9 was tested for an I1 mutation and was negative but he was not tested for any I2(including all subclades) mutations.
There are many Y DNA I2a1 subclades in Europe today, it's most diverse in western Europe and all are probably native(Mesolithic) to western Europe. I2a1b1 L621 is very popular in south-eastern Europe but i think is descended of post Mesolithic west Europeans. I2a2 and I1 all probably descend from central-west Mesolithic Europeans. I1(which has 25 defining mutations) may not have existed in the Mesolithic but it's ancestral version certainly did, probably somewhere in central Europe. I2a2 was largely dispersed by Celts and Germans, but it did exist somewhere in Mesolithic central-west Europe. For more about Y DNA I today: Eupedia Y DNA I1, Eupedia Y DNA I2
Pigmentation


On the right is a reconstruction of 7940-7690BP Mesolithic European(northern Spain) hunter gatherer named La Brana-1. On the left is a reconstruction of an ~5,300BP Late Neolithic European farmer(Alps) named Otzi based. The reconstructions are based on their skeletal remains and alleles in SNP's associated with pigmentation.
On the right is a reconstruction of 7940-7690BP Mesolithic European(northern Spain) hunter gatherer named La Brana-1. On the left is a reconstruction of an ~5,300BP Late Neolithic European farmer(Alps) named Otzi based. The reconstructions are based on their skeletal remains and alleles in SNP's associated with pigmentation.
New SNP's associated with pigmentation were tested for Loschbour, Stuttgart, and Motala12(Motala12 was not tested at all in their previous preprint). The same 8plex and Hirisplex were listed for Loschbour and Stuttgart, but some results were also listed for Motala12.
Here are the predictions for hair and eye color of Loschbour and Stuttgart using the Hirisplex model.
| Hair color | Loschbour | Stuttgart |
| Brown | 41.30% | 22% |
| Red | 0% | 0% |
| Black | 57.90% | 77.40% |
| Blond | 0.80% | 0.50% |
| Hair shade | Loschbour | Stuttgart |
| Light | 2.20% | 0.60% |
| Dark | 97.80% | 99.40% |
| Eye color | Loschbour | Stuttgart |
| Blue | 61.30% | 0% |
| Intermediate | 16.60% | 0.40% |
| Brown | 22.20% | 99.60% |
Motala12(Sweden, 6,000BC) like Loschbour(Luxembourg, 6220-5990 BC) and La brana-1(Spain, 5940-5690BC) most likely had light(prob. blue) eyes and dark hair. That's 3/3 for Mesolithic Europeans which means that combination was widespread and very popular in Europe by at least 8,000 years ago. The most compelling evidence Motala12 also had light eyes(probably blue) is that he had G/G alleles in SNP rs12913832.
Motala12 had T/T alleles in SNP rs12203592(in gene IRF4) like Loschbour and La Brana-1(T/C). Today T/T and T/C alleles in that SNP are very rare in modern Europeans and absent in east asians and Africans(see here). IRF4 is associated with sensitivity of skin to sun exposure, freckles, blue eyes, and brown hair color(got that from here).
Alot of the stuff about pigmentation is new to me so I am going to give some quotes.
We typed the three ancient modern humans at 7 SNPs forming three short haplotypes associated with eye color in present-day worldwide populations (Table S8.3)26. The observed reads in the Motala12 forager, like the Loschbour forager match the blue-eye-associated allele at all 7 SNPs.
We find that the Loschbour forager is homozygous for the h-1 HERC2/OCA2 haplotype observed in 97% of blue-eyed individuals in a present-day study population from Turkey, Jordan, and Denmark.
Motala12 was tested for 10 of 13 H-1 HERC2/OCA2 haplotype SNPs and had all of them except in rs3935591 he had T/C. 97% of modern blue eyed people(and Loschbour) have C/C. Of the mutations tested and associated with blonde and red hair none of the ancient samples had any.
Motala12 surprisingly had alleles A/A('light skin" version) in SNP rs1426654(in gene SLC24A5) like near eastern(main ancestors migrated to Europe) farmers Stuttgart and Otzi and unlike fellow European hunter gatherers Loschbour and La Brana-1 who had G/G.
There is another similarity between Motala12, near eastern farmers, and most modern west Eurasians with SNP's associated with skin color.
Examining the SLC24A5 region, we find that the Stuttgart farmer is homozygous for the C11 haplotype found in 97% of all modern carriers of the derived rs1426654 pigmentation-lightening allele27. The A111T mutation is estimated to have arisen at ~22-28 kya28, with the selective sweep favoring its rise beginning ~19kya (under a dominant model) or ~11kya (under an additive model)29. The Loschbour forager does not carry the derived rs1426654 allele. The Motala12 forager, like the Stuttgart farmer, is homozygous for the C11 haplotype.
This means that middle eastern ancestors are not the sole source of the derived rs1426654 pigmentation-lightening allele in modern Europeans. My opinion is that the skin color of these ancient hunter gatherers is unknown. A good guess is that they had dark skin, but there is evidence in modern people that they had light skin and possibly some(in eastern Europe?) who have not been sampled had blonde and red hair. The farmers were probably as light as Sardinians(their closest relatives), as dark as west Asians, or somewhere in-between. It is pretty certain that both European hunter gatherers and near eastern farmers had primarily dark hair, but the hunter gatherers were mainly light eyed while the farmers were mainly dark eyed.
Of the three mutations(in genes TYR, SLC24A5, and SLC45A2) most associated with light skin in Europe(all are just as popular in west Asians except the one in gene SLC45A2 which is ~50% in west Asians and ~100% in Europeans) La brana-1 had 0/3, Loschbour had 0/3, Motala12 had 1/3, Stuttgart had 2/3, and of tested Otzi had 2/2. Copper age(Yamna and Catacomb cultures) and Eneolithic people of the Pontiac steppe have been found to most likely have had around 90% brown eyes, an even higher percentage of dark hair, and possibly dark skin(click here). In contrast bronze and iron age Indo Iranians of south Siberia who are their supposed descendants and relatives had majority light eyes, light hair, and probably light skin(of SNP's associated with pigmentation tested their frequencies were no different from modern northern and eastern Europeans).
The distribution of light eyes and dark eyes in modern Europeans is nearly identical to the distribution of Mesolithic and near eastern ancestry, which makes sense since so far 3 out of 3 Mesolithic Europeans tested have light eyes and 2 out of 2 early European farmers tested so far have dark eyes. I think light eyes in modern European's ancestors probably went through powerful selection after the farmers and hunter gatherers mixed but the mutations for it mainly came from their hunter gatherer ancestors.
So far 3 out of 3 Mesolithic Europeans and 2 out of 2 Neolithic Europeans tested fo SNPs that can accurately determine hair color have no mutations associated with blonde or red hair. It would make sense that high percentages of light hair originated in Mesolithic Europeans since they were mainly light eyed(light haired people have a much higher percentage of light eyes). I think it is likely that light hair did exist in Mesolithic Europeans(and early farmers) just because they were west Eurasian but that doesn't mean a high percentage of them did. The origin of a high percentage of non dark hair in modern Europeans(unlike any other people in the world) is still a mystery.
Last edited: