I have revised all the
mtDNA frequencies and added 20 new populations. This now permits me to create mtDNA maps.
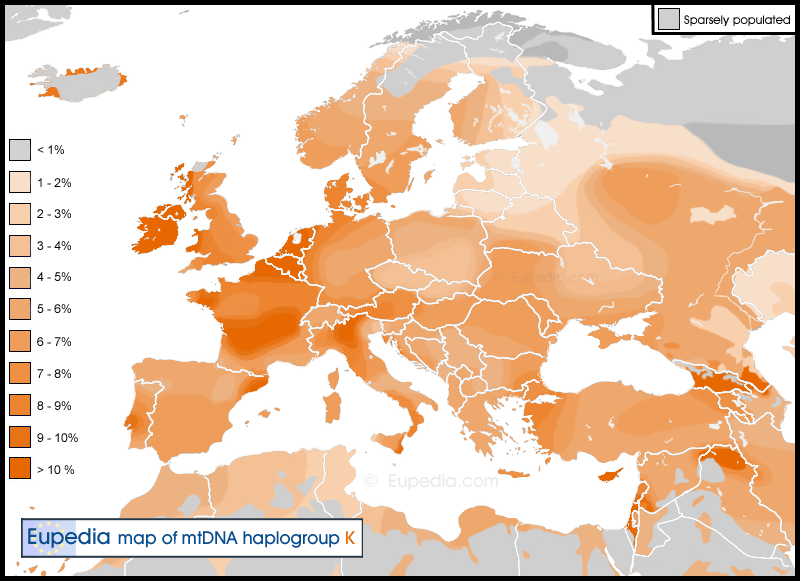
Ideally I would need more detailed regional data for central and southern Spain, all Germany, Ukraine and European Russia (except the Caucasus, which is well covered). I have got especially conflicting data for Russia, where frequencies for haplogroup K vary from 0% near Karelia to nearly 8% in nearby Vladimir and Yaroslav (around Moscow) but an average of just 3.7% for the 1768 samples collected.
The frequency of haplogroup K seems to correlate with that of haplogroup R1b in Europe (although not in the Near East and North Africa). It has been proven by ancient DNA that hg K arrived in Europe during the Neolithic and Chalcolithic, and was apparently more prevalent back then than today. So why the correlation with R1b, which only came during the Bronze Age ? I believe that this is because R1b people originated in the Near East, then picked up maternal K lineages in the Caucasus (Georgia has the highest frequency of any country), then again in Southeast Europe before migrating to Western Europe. Nowadays the highest frequencies of mtDNA K in Europe are all found in regions with high R1b levels, such as Ireland, the Western Isles of Scotland, western Wales, the Benelux, Denmark, western France, Catalonia and northern Italy (especially northern Tuscany where R1b is particularly high). The Basques are an exception, but
that was completely expected (my theory being that the Basques did not adopt an IE language because their retained essentially pre-IE maternal lineages).
All Neolithic lineages, both maternal and paternal, have declined in frequency with the arrival of the Indo-Europeans. I do not doubt that many K subclades in Europe originated with the Neolithic farmers, but they probably make up less than half of the
number of modern lineages.
An analysis of the frequencies of K subclades would allow us to determine which subclades correlate most with a dispersal from the Caucasus/Balkans by R1b Indo-Europeans. At first sight I would say that K1a4 could be one of them, because it is found in Northeast Anatolia, Georgia southern Ukraine, Hungary, Czechia and all Western Europe. It is also the most common subclade in Northwest Europe.
Neolithic lineages would probably have a greater diversity (more subclades) because they expanded earlier. However the trimming of the Neolithic/Chalcolithic population would have slowed down their diversification from the Bronze Age. In contrast, Indo-European lineages would have thrived from the Bronze Age onwards, resulting in a sudden expansion of deep subclades. This is exactly what we observe with K1a4. The subclades that remained behind in Anatolia and the Caucasus have become K1a4f. In Ukraine, the Balkans and Germany we see the appearance of K1a4a. K1a4a1 seems to have developed in Germany, probably around the same time as the branching of R1b-L11 into P312 and U106. Then, in Western Europe we witness an explosion of subclades: K1a4a1a1, K1a4a1a2, K1a4a1a3, K1a4a1b, K1a4a1c, K1a4a1c, K1a4a1d, K1a4a1e, K1a4a1f, K1a4a1g... and more subclades under those. All of them are found in Western Europe, particularly in regions with high percentages of R1b. I am not aware of any K1a4a found in Sardinia, for instance. K1a4b, K1a4c, K1a4d, and K1a4e are much more minor but most of them are found in Germany and Western Europe too. K1a4c is found in southern Italy, Greece and western/central Anatolia and could be linked to the earlier branching off of R1b-L23.
There are surely other subclades of K linked to the diffusion of R1b. Potential candidates include K1a3, K1c1 and K2a6.
Note that haplogroup K has also been found in all Asian populations where R1b is present, including the Volga-Ural, the Altai, Mongolia, Xinjiang, and most of Central Asia.