Discussion
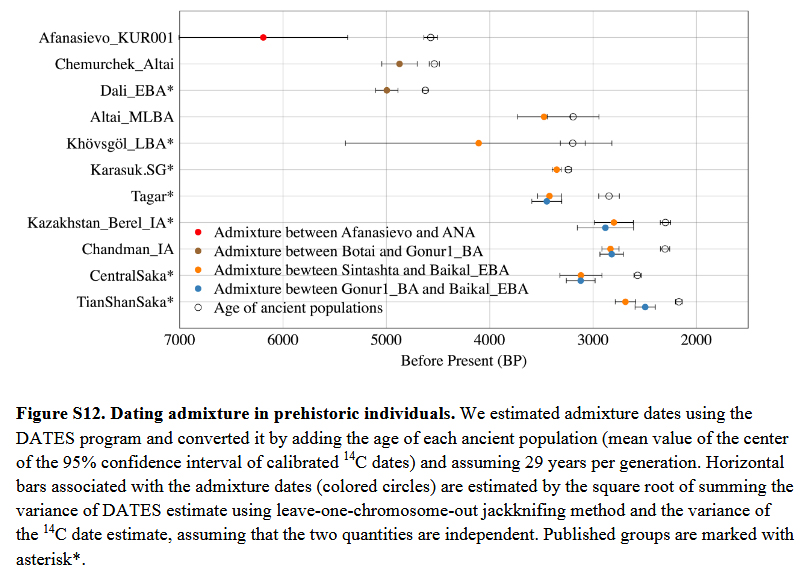
The population history of the Eastern Steppe is one marked by the repeated mixing of diverse599 eastern and western Eurasian gene pools. However, rather than simple waves of migration, demographic events on the Eastern Steppe have been complex and variable. Generating more than 200 genome-wide ancient datasets, we have presented the first genetic evidence of this dynamic population history, from ca. 4600 BCE through the end of the Mongol empire. We found that the Eastern Steppe was sparsely populated by hunter gatherers of ANA and ANE604 ancestry during the mid-Holocene, and then shifted to a dairy pastoralist economy during the Bronze Age. Migrating Yamnaya/Afanasievo steppe herders, equipped with carts and domestic606 livestock (Kovalev and Erdenebaatar, 2009), appear to have first introduced ruminant dairy pastoralism ca. 3000 BCE (Wilkin et al., 2019), but surprisingly had little lasting genetic impact, unlike in Europe (Allentoft et al., 2015; Haak et al., 2015; Mathieson et al., 2015). Subsequent Chemurchek pastoralists in the Altai were confirmed in this study to represent a separate migration of dairy pastoralists with ANE and Iranian-related ancestry, who possibly migrated into the Altai region from the south, likely via Xinjiang and/or mountainous Central Asia due to the concentration of Chemurchek burials in this region (Jia and Betts, 2010). By the MLBA, ruminant dairy pastoralism had been adopted by populations throughout the Eastern Steppe (Wilkin et al., 2019), regardless of ancestry, and this subsistence has continued, with the additions of horse milking in the LBA and camel milking in the Mongol period (Wilkin et al., 2019), to the present day (Bat-Oyun et al., 2015; Kindstedt and Ser-Od, 2019). Puzzlingly, however, there is no evidence of selection for lactase persistence over this 5,000-year history, despite the repeated introduction of this genetic trait by subsequent migrations of groups from the west. This suggests a different trajectory of lactose adaptation in Asia that to date remains unexplained. During the MLBA, we observed the formation of a tripartite genetic structure on the Eastern Steppe, characterized by the continuation of pre-Bronze ANA ancestry in the east and a cline of genetic variation between pre-Bronze Age ANA-ANE ancestry in the north and increasing proportions of a new Sintashta-related WSH ancestry in the west. The Sintashta, a western forest steppe culture with genetic links to the European Corded Ware cultures (Mathieson et al., 2015), were masters of bronze metallurgy and chariotry (Anthony, 2010), and the appearance of this ancestry on the Eastern Steppe may be linked to the introduction of new (especially horse-related) technologies. DSKC sites in particular show widespread evidence for horse use in transport, and perhaps even riding (Taylor et al. 2015), and genetic analysis has demonstrated a close link between these animals and the Sintashta chariot horses (Fages et al., 2019). The strong east-west genetic division among Bronze Age Eastern Steppe populations at this time was maintained for more than a millennium and through the end of the EIA, when the first clear evidence for widespread horseback riding appears (Drews, 2004) and the heightened mobility of some groups, notably the eastern Slab Grave culture (Honeychurch, 2015), began to disrupt this structure. Eventually, the three major ancestries met and mixed, and this was contemporaneous with the emergence of the Xiongnu empire. The Xiongnu are characterized by extreme levels of genetic heterogeneity and increased diversity as new and additional ancestries from China, Central Asia and the Western Steppe (Sarmatian-related) rapidly entered the gene pool. Genetic data for the subsequent early medieval period are relatively sparse and uneven, and few Xianbei or Rouran sites have yet been identified during the 400-year gap between the Xiongnu and Türkic periods. We observed high genetic heterogeneity and diversity during the645 Türkic and Uyghur periods, with slight shifts in their Sarmatian-like western ancestry component towards that seen among the contemporaneous Alans, a Sarmatian-descendent group known for invading the Roman empire and warring with the Germanic tribes of Europe (Bachrach, 1973). Following the collapse of the Uyghur empire, we documented a final major genetic shift during the late medieval period towards greater eastern Eurasian ancestry, which is consistent with historically documented expansions of Tungusic- (Jurchen) and Mongolic (Khitan and Mongol) speaking groups from the northeast into the Eastern Steppe (Biran, 2012). We also observed that this East Asian-related ancestry was brought into the Late Medieval populations more by male than female ancestors, and we observed a corresponding increase in Y-haplogroups associated with southeast Asians and the purported patriline of Ghenghis Khan (O2a and C2b, respectively). By the end of the Mongol period the genetic make-up of the Eastern Steppe had dramatically changed, retaining little of the ANE ancestry that had been a prominent feature during its prehistory. Today, ANE ancestry survives in appreciable amounts only in isolated Siberian groups and among the indigenous peoples of the Americas (Jeong et al., 2019). The genetic profile of the historic Mongols is still reflected among contemporary Mongolians, suggesting a relative stability of this gene pool over the last ~700 years. Having documented key periods of genetic shifts in the Eastern steppe, future work may be able to explore whether these shifts are also linked to cultural and technological innovations and how these innovations may have influenced the political landscape. Integrating these findings with research on changes in horse technology and herding practices, as well as shifts in livestock traits and breeds, may prove particularly illuminating. This study represents the first large-scale paleogenomic investigation of the eastern Eurasian Steppe and it sheds light on the remarkably complex and dynamic genetic diversity of the region. Despite this progress, there is still a great need for further genetic research in central and eastern Eurasia, and particularly in northeastern China, the Tarim Basin, and the eastern Kazakh steppe, in order to fully reveal the population history of the Eurasian Steppe and its pivotal role in world prehistory.