
"To understand the full extent of the role played by demic processes in this key transition in Late Glacial Europe we focused on the left hemimandible of an individual found at Riparo Tagliente (Tagliente2) associated with Late Epigravettian evidence. During the LGM and the Late Glacial, the Adriatic Sea basin played a critical role in shaping the economy and mobility of Epigravettian groups. Geomorphological and sedimentological processes linked to the extension of Alpine glaciers and to the lowering of the sea level at ~120m amsl formed a vast plain area that connected the Italian and Balkan peninsulas. The central Slovenian passage was used by large pachyderms and ruminants such as cervids and bisons which, alongside with bears, were the main game targeted by Epigravettian hunter gatherers. Riparo Tagliente (northeastern Italy) represents the earliest available evidence of human occupation of the southern Alpine slope while the main glaciers in the area started withdrawing 17.7 ka ago, and is therefore critical to address questions on the impact of human movement in this time frame.
Y-DNA I2-M436, mtDNA U4’9
Overall contamination estimated from mtDNA was 2.158% and 0.60 – 1.53% from the X chromosome. The mtDNA haplogroup is a basal U4’9, consistent with a European Palaeolithic individual.
X/Autosome coverage ratio in the order of 0.56 confirmed the individual was male, and the chromosome Y haplogroup estimated to be I2, which captures the majority of the post-Villabruna diversity in Europe. "
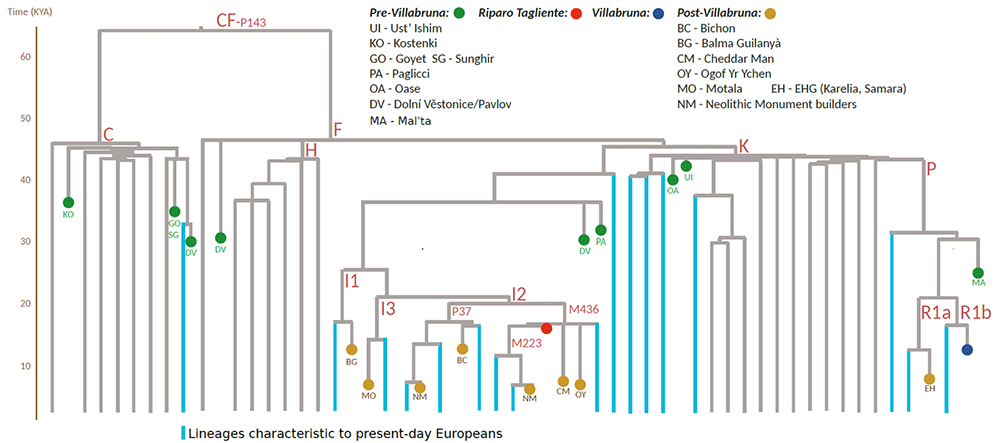
( ChrY haplogroup of Tagliente2 (in red) surrounded by post-Villabruna samples (including Bichon, BC). Y Haplogroup splits are drawn according to the dater estimates based on high coverage modern sequences from Karmin et al. 2015. The ancient individuals are mapped on this tree considering the available haplogroup-informative available SNP data and private mutations in the ancient samples have been ignored. )
Western European Hunter Gatherer
From a population perspective, we performed a MDS analysis based on outgroup f3 distances and found the sample to fall within the broader European Western Hunter Gatherer (WHG) genetic variation, pointing to an affinity to the previously described Villabruna Cluster, known to have largely replaced previous European Hunter Gatherer populations at least ~14 ky ago. One of the defining feature of the Villabruna cluster is a higher affinity with Near Eastern genetic components, compared with pre-existing palaeolithic West Eurasians: the significantly negative f4 test (Kostenki14, Tagliente2, Druze, Mbuti: f4=-0.0037; standard error= 0.00063; Z=-5.88) further confirmed Tagliente2 to share genetic features with the Villabruna cluster and to be in discontinuity with the preceding European genetic background.
The higher affinity of Continenza and Bichon to later WHG (Loschbour, Iberia_HG and Continenza, Bichon, and Villabruna themselves) when compared to Tagliente2 may be explained by the more ancient age of Tagliente2 or with the former individuals being genetically closer to the genetic ancestry that reached central Europe at least by 14ka ago. Alternatively, the higher affinity emerging among more recent WHG samples may also be ascribed to subsequent admixtures between the newly arrived Tagliente2 individuals and preexisting Dolní Věstonice- or Goyet-like (~35ka ago) genetic substrates, as already reported for Loschbour (~8.1ka ago). (…)
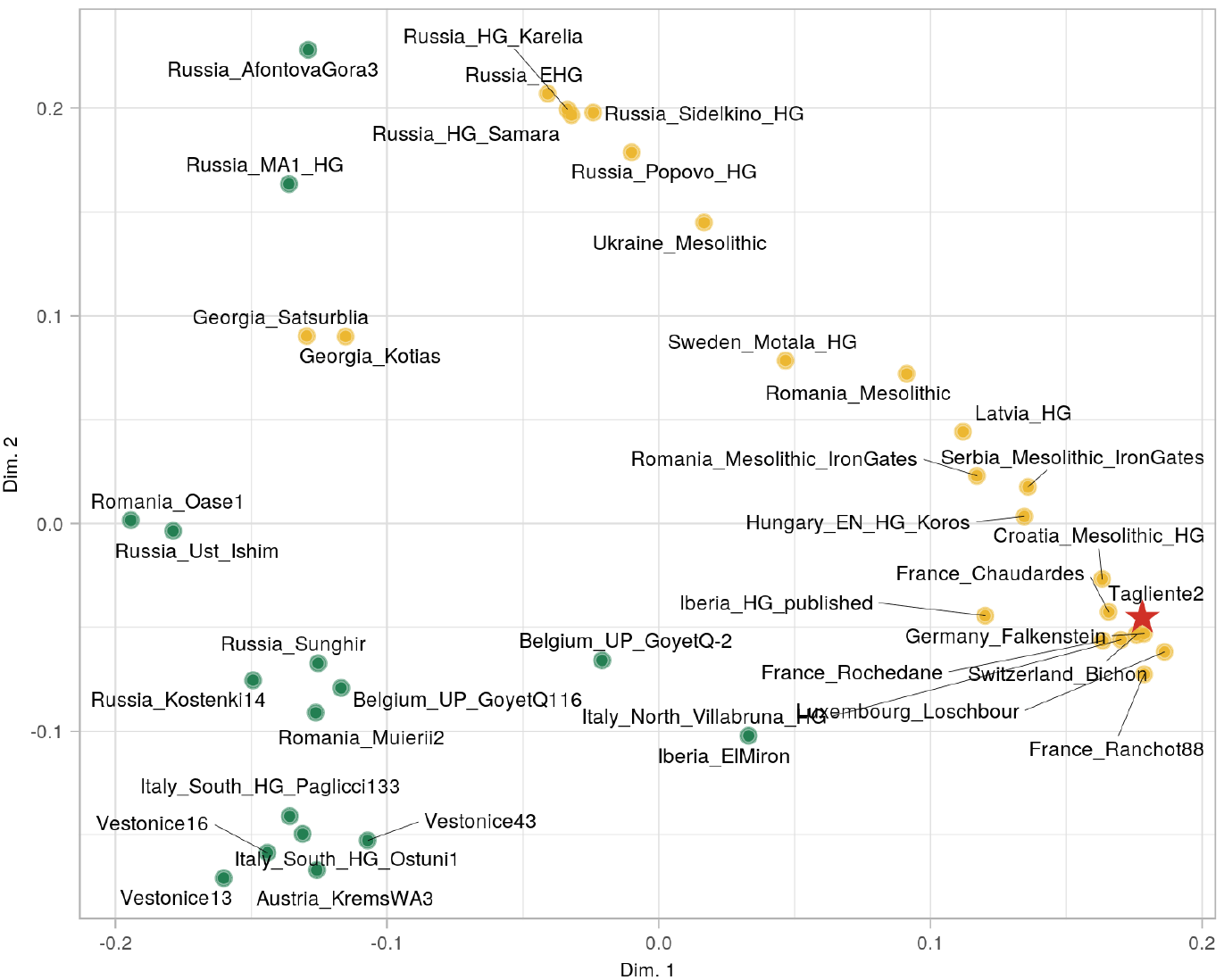
( Demographic inference from Tagliente 2. Mutidimensional Scaling based on (Mbuti; X, Tagliente2) outgroup f3 statistics show Tagliente2 (red star) to cluster within the Villabruna Cluster (in gold) and away from the pre-existing South European samples (in green).
Evidence of earlier migrations
Tagliente2 therefore provides evidence that the major migrations which strongly affected the genetic background of all Europeans, started considerably earlier in Southern Europe than previously reported, and in this region they do not seem to be limited to favourable, warmer periods (e.g. Greenland Interstadial 1, ~14.7-13ka ago). Our results rather show that population movements were already in place during the cold phase immediately following the LGM peak. At this stage, Italy, the Balkans, and Eastern Europe/Western Asia were already connected into the same network of potential LGM refugia, and exchanged both genes and cultural information.
This finding also backdates previous conclusions concerning a plausible demic component to change over time in the coeval material culture of Southern Europe, and temporally locates this process at the transition between Early and Late Epigravettian or even possibly at an early stage of the Epigravettian and at the very beginning of the Magdalenian sequence. The latter scenario stems from interpreting the traces of Villabruna/Tagliente2 genetic components recorded at El Mirón (~19ka ago) as the result of an early phase of this westward expansion, rather than as a proliferation from Western European refugia. Further genetic evidence from Southern European contexts dated between ~24-19 ka ago, however, will be needed in the future to test this hypothesis.
(…) Tagliente2, therefore, suggests that cumulative cultural change observed in Southern Europe from the end of LGM to the end of the Younger Dryas (~11.7 ka ago) was at least in part triggered by gene flow from eastern refugia into Northeastern Italy and that this process, in its early stage, was independent of warming events, and contributed to the gradual replacement of pre-LGM ancestry across the Italian peninsula.
Source: https://www.biorxiv.org/content/10.1101/2020.08.10.241430v1
Y-DNA I2-M436, mtDNA U4’9
Overall contamination estimated from mtDNA was 2.158% and 0.60 – 1.53% from the X chromosome. The mtDNA haplogroup is a basal U4’9, consistent with a European Palaeolithic individual.
X/Autosome coverage ratio in the order of 0.56 confirmed the individual was male, and the chromosome Y haplogroup estimated to be I2, which captures the majority of the post-Villabruna diversity in Europe. "
( ChrY haplogroup of Tagliente2 (in red) surrounded by post-Villabruna samples (including Bichon, BC). Y Haplogroup splits are drawn according to the dater estimates based on high coverage modern sequences from Karmin et al. 2015. The ancient individuals are mapped on this tree considering the available haplogroup-informative available SNP data and private mutations in the ancient samples have been ignored. )
Western European Hunter Gatherer
From a population perspective, we performed a MDS analysis based on outgroup f3 distances and found the sample to fall within the broader European Western Hunter Gatherer (WHG) genetic variation, pointing to an affinity to the previously described Villabruna Cluster, known to have largely replaced previous European Hunter Gatherer populations at least ~14 ky ago. One of the defining feature of the Villabruna cluster is a higher affinity with Near Eastern genetic components, compared with pre-existing palaeolithic West Eurasians: the significantly negative f4 test (Kostenki14, Tagliente2, Druze, Mbuti: f4=-0.0037; standard error= 0.00063; Z=-5.88) further confirmed Tagliente2 to share genetic features with the Villabruna cluster and to be in discontinuity with the preceding European genetic background.
The higher affinity of Continenza and Bichon to later WHG (Loschbour, Iberia_HG and Continenza, Bichon, and Villabruna themselves) when compared to Tagliente2 may be explained by the more ancient age of Tagliente2 or with the former individuals being genetically closer to the genetic ancestry that reached central Europe at least by 14ka ago. Alternatively, the higher affinity emerging among more recent WHG samples may also be ascribed to subsequent admixtures between the newly arrived Tagliente2 individuals and preexisting Dolní Věstonice- or Goyet-like (~35ka ago) genetic substrates, as already reported for Loschbour (~8.1ka ago). (…)
( Demographic inference from Tagliente 2. Mutidimensional Scaling based on (Mbuti; X, Tagliente2) outgroup f3 statistics show Tagliente2 (red star) to cluster within the Villabruna Cluster (in gold) and away from the pre-existing South European samples (in green).
Evidence of earlier migrations
Tagliente2 therefore provides evidence that the major migrations which strongly affected the genetic background of all Europeans, started considerably earlier in Southern Europe than previously reported, and in this region they do not seem to be limited to favourable, warmer periods (e.g. Greenland Interstadial 1, ~14.7-13ka ago). Our results rather show that population movements were already in place during the cold phase immediately following the LGM peak. At this stage, Italy, the Balkans, and Eastern Europe/Western Asia were already connected into the same network of potential LGM refugia, and exchanged both genes and cultural information.
This finding also backdates previous conclusions concerning a plausible demic component to change over time in the coeval material culture of Southern Europe, and temporally locates this process at the transition between Early and Late Epigravettian or even possibly at an early stage of the Epigravettian and at the very beginning of the Magdalenian sequence. The latter scenario stems from interpreting the traces of Villabruna/Tagliente2 genetic components recorded at El Mirón (~19ka ago) as the result of an early phase of this westward expansion, rather than as a proliferation from Western European refugia. Further genetic evidence from Southern European contexts dated between ~24-19 ka ago, however, will be needed in the future to test this hypothesis.
(…) Tagliente2, therefore, suggests that cumulative cultural change observed in Southern Europe from the end of LGM to the end of the Younger Dryas (~11.7 ka ago) was at least in part triggered by gene flow from eastern refugia into Northeastern Italy and that this process, in its early stage, was independent of warming events, and contributed to the gradual replacement of pre-LGM ancestry across the Italian peninsula.
Source: https://www.biorxiv.org/content/10.1101/2020.08.10.241430v1