We might deal with not just one but multiple mixture events. Like e.g. the initial spread of the CHG component along the coast with those fisher-pastoralist population which reached the Lower Don might have been dominated by CHG on both the male and the female side, but male totally. They could have incorporated local EHG females (no or very few males) and this mixed population with say 75 : 25 (CHG : EHG) could then have been taken over by male foragers from the North.
Having multiple mixture event didn't saves the day for a male-EHG dominated build-up of the WSH signal.
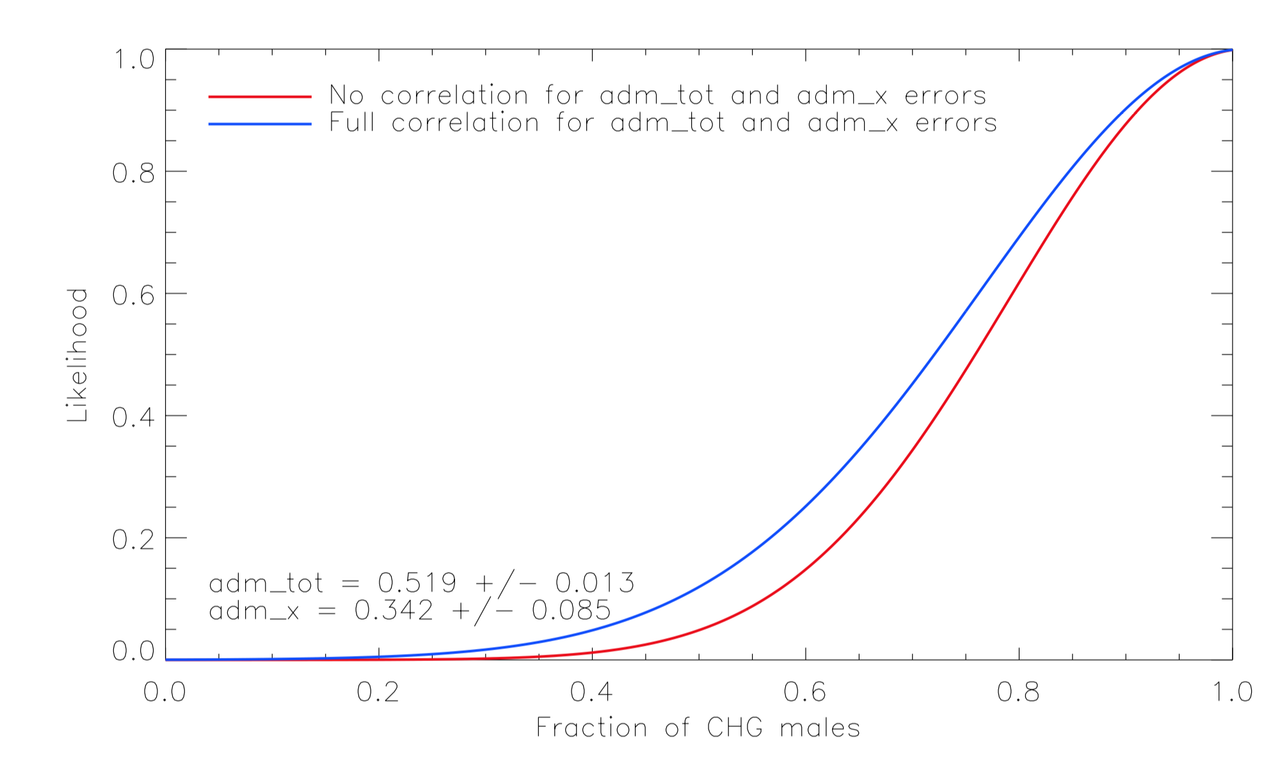
For instance, in a single admixture model, here is the likelihood according to Lazaridis numbers (indeed, not strong constraints, but still definitely in favor of CHG-males) :
I computed the constraints under two extreme hypotheses :
(i) The errors are dominated by the limited amount of data and thus nearly uncorrelated.
(ii) The errors are "model dominated" and therefore ~fully correlated.
It didn't make a big difference anyway.
At least in such setting it shows how disfavored is a male-EHG bias.
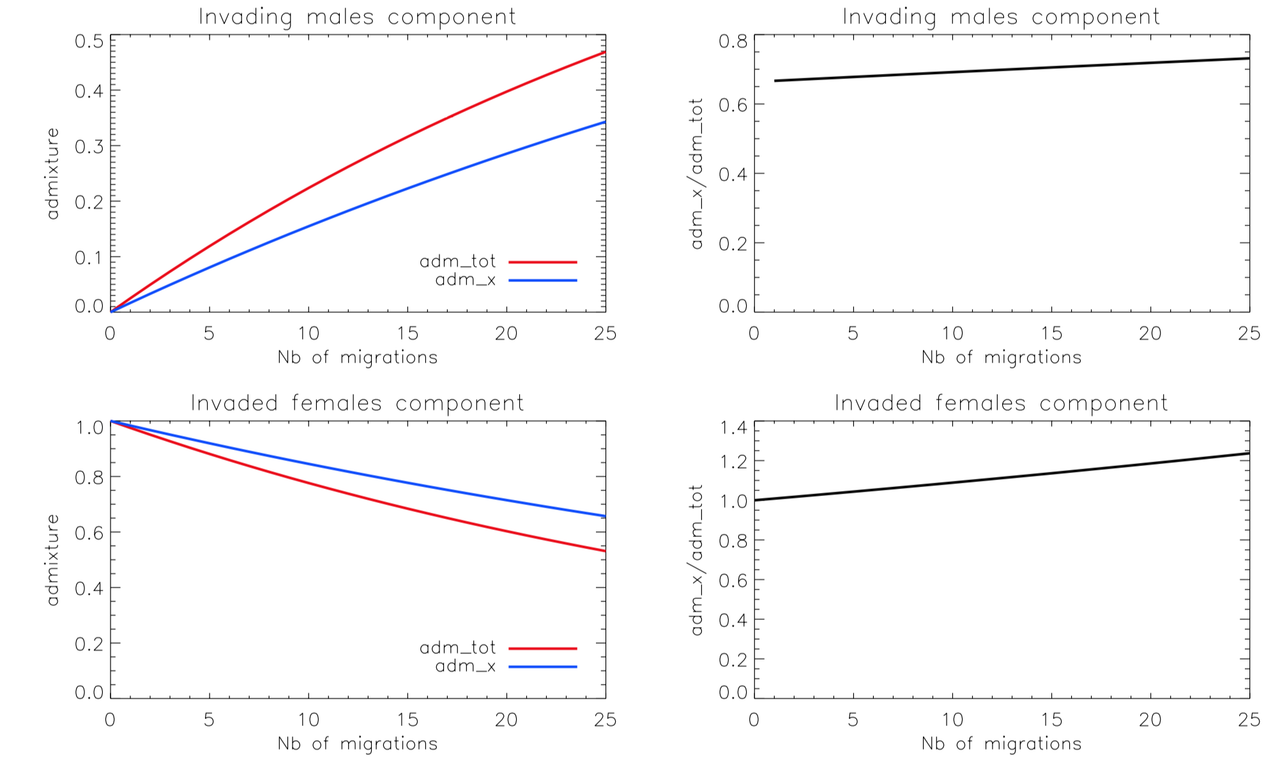
Then we can look at a smooth admixture, with "new comers" each generations :
As we see, in such scenario, the X chromosome (adm_x) still receives higher contribution from the female source populations.
Therefore, if you merge EHG-males and CHG/EHG admixed women, chrX will have less EHG than CHG.
Such scenario fails to produce a boost for the sex bias.
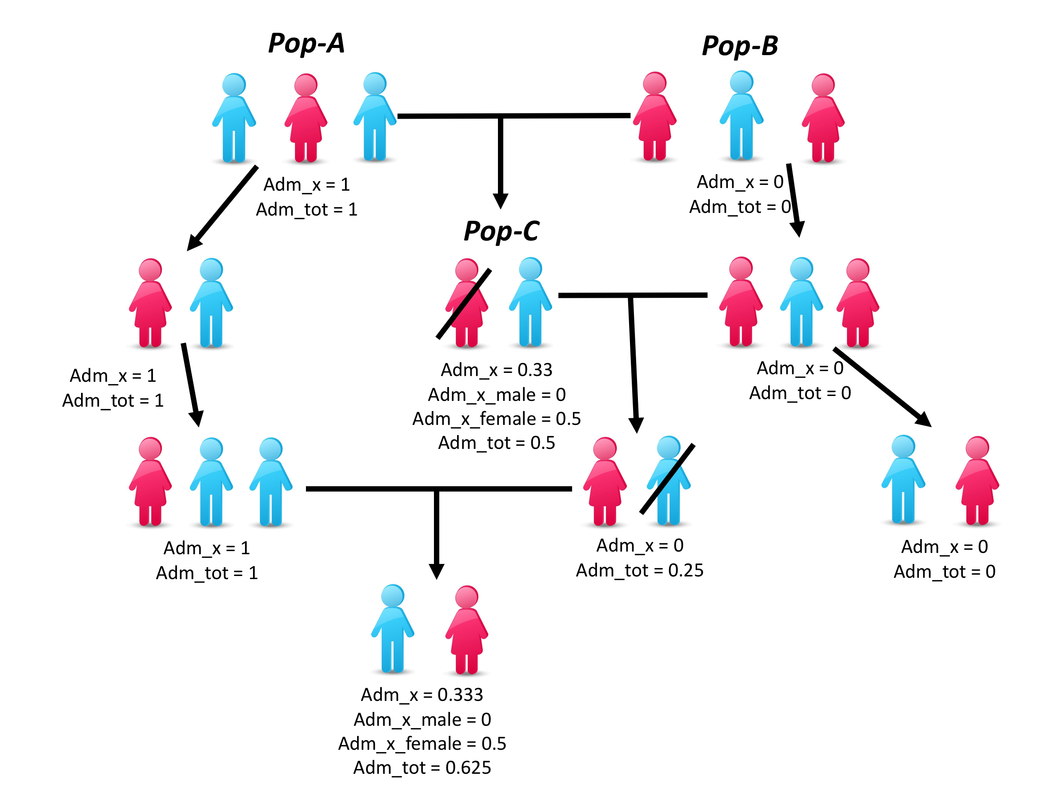
However, there is an "easy" way to "enhance" the sex bias derived from X-chromosome admixture, which consits in a sequenced patern of reproduction between two source populations progressively merging into a third new population, here is a small schema (each arrow represent only one generation, and a black line on some individual indicates that they got replaced or excluded from reproduction) :
This mixture scenario "takes advantage" of the fact that first-gen admixed males (in a heavily sex-biased merging scenario) only have admixture from the source population of the females on the X chromosome.
Then, by avoiding first-gen admixed females to participate to the reproduction, you prevent the X chromosome from the male source population to be passed down.
By having the first-gen admixed male mating with female's source population only, you have "cleaned" all contribution on the X chromosome from the male source population. However, you drop at only ~25% of total autosomal contribution for the male source population after two generations.
Then, you can increase again this component with a new wave of males from the male source population.
If you repeat this patern, technically you can have after few centuries a population that will be fully sourced by the male side for all chromosomes but X and fully sourced by the female-side for the X chromosomes.
But as you see, this "bias enhancement" scenario didn't help here, as it "increase" the sex bias ... but it is completely unable to "reverse" the apparent sex-bias.
To my knowledge, you have no way to produce a "reversed" sex-bias.
I don't see any merging pattern that would be able to produce more male-side admixture on the X chromosome (you can do the opposite because you can "flood" males X with females Xs).
(PS: unless you completely replace EHGs X-chromosome with the above scenario by CHGs X-chromosomes, and at the same time you also replace the CHG-X chromosome by an EHG-X chromosome and then you'll be able to reproduced the observed numbers ... but the above scenario is already convoluted, and that would start to become "cumbersome".)
Therefore, whenever you replace CHG males by EHG males (carrying EHG autosomal DNA), you will end up with less EHG on the X chromosome, which is not what is observed.
If we accept the numbers from Lazaridis, the predominance of some Y-lineages have to be related with a later founder effects that occured post-merging.
For instance, R-M269 only started to expand around 4400 BCE, therefore, if the merging creating WSH occured in a small population around 4500 BCE, then it is fairly easy for a random lineage to take over when the population will expand.
Let imagine that the Y-lineages of the WSH post-merging were ~80% CHG and ~20% EHG (completely inside the error bars of Lazaridis numbers), then there is ~20% chances that a founder effect will concern an EHG lineage ... not the most likely, but ~20% probability is not really disturbing.
R-M417 is definitely no big-deal in such scenario, as it could be a lonely lineage that got absorbed and "demographicly flooded" by WSH before expanding around ~3400 BCE. It didn't need to have been part of the WSH population since "founding".
If the above scenario is correct, with a WSH "creation" around 4500 BCE and R-M269 becoming dominant inside a small population arond 4400 BCE, then it might take a long time before any heavily-CHG group related to the root of WSH will be found among ancient DNA.
Also, it is interesting to note that some Y-lineages with a likely origin among CHG did survived, for instance the well known J-CTS1026. Another exemple could be J-Y167175.