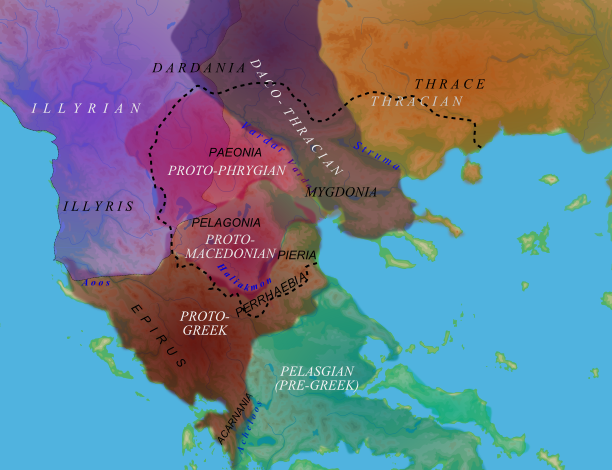
I think too the 'pelasgian' label doesn't tell us a lot... as very often it is a exogamic name
concerning Y-E-V13 I have not too much to add to what was said - someones here put solid arguments and we have now to wait for a chanceous ancient DNA in the right places and times -
here under a discussion I found in the forum "For what they were we are" - maybe I have red to quickly but I didn't see Y-DNA SNPs in it - but could it explain an old BUT neolithic presence of y-E1b and subclades in Europe??? just to feed the "war" (keep cool, it's my aperitive hour)
There are severalinteresting studies in my "to do" list and I will becommenting them in the following days (I am quite busy these weeksand therefore I concentrate my efforts on weekends).
In this entry we have arather interesting analysis of ancient mtDNA from the Pre-PotteryNeolithic B of Syria (NE and South) and its legacy on modernpopulations of West Asia and SE Europe, as well as on ancientEuropean Neolithic ones.
Eva Fernández et al.,Ancient DNA Analysis of 8000 B.C. Near Eastern FarmersSupports an Early Neolithic Pioneer Maritime Colonization of MainlandEurope through Cyprus and the Aegean Islands. PLoSGenetics 2014. Open access→ LINK[doi:10.1371/journal.pgen.1004401]
I understand that thesequences are not really new but that they were first discussed in
Fernández 2005 (thesisin Spanish) and
2008.What is new is the comparison with ancient and modern populations insearch of their possible legacy.
[TABLE="width: 322"]
[TR]
[TD="width: 322"]

[/TD]
[/TR]
[TR]
[TD="width: 322"] Early PPNB (from
CONTEXT C14 database)
[/TD]
[/TR]
[/TABLE]
In spite of the relevanceof this analysis, it must be cautioned tha
PPNBancient mtDNA and its legacyt the Tell Ramad and Tell Halulasites may not be fully representative of the actual genetic diversityof PPNB as a whole, a cultural area that spanned all the Levant, fromthe Kurdish mountains to the Sinai and Cyprus.
If, as the authors argueand I have already suggested in relation to the NE African affinitiesof European Neolithic ancestry, the arrival of Neolithic to Thessalyhappened via a coastal route, inland PPNB sites may well not be asinformative as Palestinian or Cypriot ones.
But this is what we havefor now, so let's see what these ancient Syrian farmers tell us,while we await further Neolithic sequences from potentially morerelevant sites.
[TABLE="width: 602"]
[TR]
[TD="width: 602"]

[/TD]
[/TR]
[TR]
[TD="width: 602"]
Table 1. Mitochondrial DNA typing of 15 Near Eastern PPNB skeletons.
[/TD]
[/TR]
[/TABLE]
40% of the sequencesbelong to haplogroup K, a U8-derived lineage unknown in Europe beforethe Neolithic. Most of the other lineages (40%) belong to R0 but halfof them belong to R0(xHV), extremely rare in Europe (common in Arabiainstead) and the H sequences cannot be identified either withanything common nowadays. The remaining 20% of lineages (U*, N* andL3*) are not too helpful either.
So when the authorscompare them with modern and ancient populations most of the affinitycorresponds to
a single basal haplotype of K (16224C,16311C)as described in supplementary table 5.
[TABLE="width: 643"]
[TR]
[TD="width: 643"]

[/TD]
[/TR]
[TR]
[TD="width: 643"]
Figure 2. Contour map displaying the percentage of individuals of the database carrying PPNB haplotypes.
Only populations with clear geographic distribution were included. Gradients indicate the degree of similarity between PPNB and modern populations (dark: high; clear: small).
[/TD]
[/TR]
[/TABLE]
The SE European and WestAsian populations with the greatest legacy of this haplotype are: the
Csángó ofMoldavia (22%), Cypriots (13%), Ashkenazi Jews (11%), Crimean Tatars(10%) and Georgians (9%). Cardium Pottery farmers from Catalonia(23%) and a pooled Central European Danubian Neolithic sample (10%)also score high for this lineage.
Some other PPNB matrilineages also show some lessermodern prevalence:
- 16223T (L3) → Qatar, Yemen (not necessarily the same L3(xM,N) lineage, it must be said)
- 16224C,16311C,16366T (K) → Druze
- 16256T (H) → Bedouin
The other haplotypes havenot been detected in modern nor European Neolithic populations.
The obvious conclusion isthat only the 16224C+16311C K haplotype was, of all the EuphratesPPNB lineages active in the Neolithic European founder effect. Thishaplotype was present only in 1/15 individuals from the EuphratesPPNB, so rather marginal over there, although a close relative foundtoday among the Druze was more common (3/15).
Another conclusion isthat the
Csángó probably have a quite direct line ofancestry to the early European farmers, shedding some light on theorigin of this mysterious population at risk of extinction.
The
coastal route toThessaly proposed here makes all sense to me because, on oneside, early Anatolian Neolithic cultures do not seem to have anyobvious cultural affinity with the first European Neolithic of Sesklo(Painted Pottery) and Otzaki (Cardium Pottery), and, on the otherside, there is clear evidence of some NE African genetic legacymediated by Palestine: Y-DNA E1b-V13 naturally but also the "BasalEurasian" speculation of
Lazaridisthat ended up being revealed as Dinka affinity in fig. S7 of
Skoglund& Malström.
This theory can only bestrongly confirmed if Palestinian and Cypriot ancient DNA issequenced and fits well in it. Similarly ancient Balcanic DNA wouldbe most interesting to have as well for a more direct reference. But,in any case, the theory seems at the very least plausible andsupported by some important evidence.
[TABLE="width: 643"]
[TR]
[TD="width: 643"]
[/TD]
[/TR]
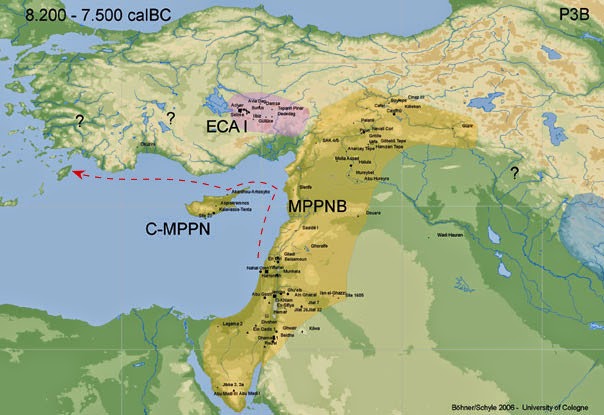
[TR]
[TD="width: 643"] My hypothetical reconstruction of a plausible coastal route of Neolithic towards Thessaly
(dashed red line)
on a base map of Middle PPNB
from the CONTEXT database.
[/TD]
[/TR]
[/TABLE]
It is also important tonotice that the Syrian PPNB sequences are different from the modernmtDNA pool of West Asia, dominated by lineages like J, T1 and U3.This suggests that, at the very least in this region of the SyrianEuphrates, there have been important demographic changes sinceNeolithic, something confirmed by data from the same are but of laterdates (which anyhow is not yet modern either).
Fernández et al. discuss this issue in some detail:
Our PPNB population includes a high percentage(80%) of lineages with a Palaeolithic coalescence age (K, R0 and U*)and differs from the current populations from the same area, whichexhibit a high frequency of mitochondrial haplogroups J, T1 and U3(TableS7). The latter have been traditionallylinked with the Neolithic expansion due to their younger coalescenceage, diversity and geographic distribution [11],[12],[49].In addition to the PPNB population, haplogroup T1 is also absent inother Early Neolithic populations analyzed so far [17],[22],[26],[30].Haplogroup U3 has been found only in one LBK individual and it hasbeen suggested that it could have been already part of thepre-Neolithic Central European mitochondrial background[19].
HaplogroupJ is present in moderate frequencies in Central European LBK-AVKpopulations (11.75%) and it has been proposed as part of the CentralEuropean “mitochondrial Neolithic package” [19].However, it has also been described in one late hunter-gathererspecimen of Germany, raising the possibility of a pre-Neolithicorigin [23].Haplogroup J is present in low frequency (4%) in Cardial/EpicardialNeolithic samples of North Eastern Spain [27],[28],[31].Absence of Mesolithic samples from the same region prevents makingany inference about its emergence during the Mesolithic or theNeolithic. However, its absence in the PPNB genetic backgroundreinforces the first hypothesis.
Thesefindings suggest that (1) late Neolithic or post-Neolithicdemographic processes rather than the original Neolithic expansionmight have been responsible for the current distribution ofmitochondrial haplogroups J, T1 and U3 in Europe and the Near Eastand (2) lineages with Late Paleolithic coalescent times might haveplayed an important role in the Neolithic expansive process. Thefirst suggestion alerts against the use of modern Near Easternpopulations as representative of the genetic stock of the firstNeolithic farmers while the second will be explored in depth in thefollowing section.
From the viewpoint ofmaterial Prehistory, it is of course correct, that PPNB wasoverwhelmed by later cultural processes, which may have implied demicexpansions and replacements of some sort, even if many of them seemto originate within West Asia.
First of all, there isthe Halafian cultural expansion, originating in Upper Mesopotamia;then we also have to consider the Semitic cultural and linguisticexpansion, originating in Palestine; finally we have to consider theIndoeuropean waves: first the Anatolian group (Hittites, etc.) viathe Caucasus, later the Balcanic group of Phrygians (and probablyArmenians as derived branch) and finally the Iranian one from CentralAsia. Even within the Semitic expansion there were probably severalwaves as well. All together must have significantly reshuffled thegenetic landscape of the region.
But unless we get moreancient West Asian DNA it will be most difficult to discern clearlyhow all that played out. After all the Syrian Euphrates can beexceptional in many aspects, being right in the middle of all: a truepivot of the Fertile Crescent, subject to pressure from alldirections.
AT FIRST SIGHT it doesn' t fit the maximum of diversity of E1-V13 (a spot in Montenegro)
I'll try to think again about it but...