
Fire Haired14
Banned
- Messages
- 2,185
- Reaction score
- 582
- Points
- 0
- Y-DNA haplogroup
- R1b DF27*
- mtDNA haplogroup
- U5b2a2b1
Today(April 24, 2014) possibly the greatest ancient DNA paper since Lazaridis 2014 'appeared in Science" Skoglund et al. 2014. I was informed by More ancient Scandinavians (Skoglund, Malmström et al. 2014). I just want to make people aware, i will add my own opinion after reading its Supplementary Materials.
There is already quite a collection of stone age Swedish hunter gatherer and farmer DNA. The individuals from this study are highlighted. If individuals in older studies were sampled from the same site as samples in Skoglund et al. 2014 but were given a differnt date, I replaced the old date with the one given in this paper. Ancestral Journeys.org is my source for the stone age Swedish individuals not in Skoglund et al. 2014
Mesolithic Swedish hunter gatherersStoraFörvar11 aka SfF11(Male), 7,500-7,250 cal. B.P, Stora Karlso Sweden : mtDNA=U5a1
6,873 ± 119 BC, Stora Karlso Sweden : mtDNA=U4b1
Motala1(Female), 6,000BC Motala Sweden: mtDNA=U5a1
Motala2(Male), 6,000BC Motala Sweden: Y DNA=I* (I P38+, I PF3742+, I L41+, I1 S108-, I1 L845-, I1 M253-, I2a1b CT1293-, I2a2 L37-), mtDNA=U2e1
Motala3(Male) 6,000BC Motala Sweden: Y DNA=I2a1b*(I M258+, I PF3742+, I2 L68+, I2a1 P37.2+, I2a1b CTS7218+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b1 M359.2-, I2a1b3 L621-), mtDNA=U5a1
Motala4(Female) 6,000BC Motala Sweden: mtDNA=U5a2d
Motala6(Male) 6,000BC Motala Sweden: Y DNA=? (Q1 L232- Q1a2a L55+), mtDNA=U5a2d
Motala9(Male) 6,000BC Motala Sweden: Y DNA=I* (I P38+, I1 P40-), mtDNA=U5a2
Motala12(Male) 6,000BC Motala Sweden: Y DNA=pre-I2a1b or brother lineage to I2a1b(I PF3742+, I M258+, I M170+, I2 L68+, I2a L460+, I2a1 P37.2+, I2a1b CTS7218+, I2a1b CTS5985+. I2a1b L178+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b CTS5375-, I2a1b CTS8486-, I2a1b1 M359.2-, I2a1b3 L621-), mtDNA=U2e1
Neolithic Swedish hunter gatherers of the Pitted Ware culture
Ajv52A(Male), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=V
Ajv59(Male), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U
Ajv53(Female), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4d
Ajv58(Male), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: Y DNA=I2a1-P37.2, mtDNA=U4d
Ajv70(Male), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4d
Ire8(Male), 5,100-4,150 cal. B.P, Ire, Hangvar, Gotland Sweden: mtDNA=U4d
Ajv13(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4
Ajv52b(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4
Ajv66(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4
Ajv54(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U5
Ajv36(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U5
Ajv5(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U5a
Ajv29a(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U5a
Fir15(?), 2800-2000 BC,Fridtorp, Västerhejde, Gotland: mtDNA=U4
Fir22(?), 2800-2000 BC,Fridtorp, Västerhejde, Gotland: mtDNA=U4
Fir4(?), 2800-2000 BC,Fridtorp, Västerhejde, Gotland: mtDNA=U5
Fir27(?), 2800-2000 BC,Fridtorp, Västerhejde, Gotland: mtDNA=U5a
Ire6b(?), 5,100-4,150 cal. B.P, Ire, Hangvar, Gotland Sweden: mtDNA=T2b
Ire9(?), 5,100-4,150 cal. B.P, Ire, Hangvar, Gotland Sweden: mtDNA=U4
Ire3(?), 5,100-4,150 cal. B.P, Ire, Hangvar, Gotland Sweden: mtDNA=U4
Gökhem4(Male), 5,050-4,750 cal. years B.P.: mtDNA=H
Gökhem2(Female), 5,050-4,750 cal. years B.P.: mtDNA=H1c
Gökhem7(Female), 5,050-4,750 cal. years B.P.: mtDNA=H24
Gökhem5(Female), 5,280-4,890 cal. B.P.: mtDNA=K1e
Ste7(Female), 5,280-4,890 cal. B.P.: mtDNA=T2b
Ste7(Female), 5,280-4,890 cal. B.P.: mtDNA=J
The mtDNA of the Stone age Swedish farmers and hunter gatherers is 100% constant with what has been found in stone age farmers and hunter gatherers from other regions of Europe. The hunter gatherers have about 100% U5, U4, and U2, and the farmers mainly have modern European-specfic subclades of haplogroups which are most diverse and originated in the middle east. Since mDNA H1 and H3 are most popular in western Europe today, i bet that Neolithic west Europeans like the ones in Sweden had a very high amount and mtDNA samples from them so far is great evidence this is true. So far all U5 samples from Swedish hunter gatherers are U5a(mainly U5a1, but also U5a2 and U5a2d), like Mesolithic Russians(specifically U5a1) and unlike Mesolithic central-west Europeans who had mainly U5b(mainly U5b2) and the only U5a subclade found are U5a2(including U5a2c3* and U5a2a). The U5a in Swedish hunter gatherers may be from east European-very ANE like ancestors,, while their Y DNA I2a1-P37.2 is probably descended of central-west European ancestors.
At least paternally modern Balts and Scandinavians don't show evidence of a high amount of Mesolithic Swedish ancestry. I2a1-P37.2 today is very very rare in Scandnavians and Balts(much of it is probably east European=specfic I2a1b1a-L147.2), there are some Scandinavian P37.2 samples in FTDNA's database who have not been tested for I2a1-P37.2 subclades i bet at least have unknown specific Scandinavian SNPs and are a brotherclade to I2a1b. Y DNA I2a1-P37.2 is likely a marker of Europeans who took refuge in south-western Europe during the last ice age, today it is most diverse in western Europe and so far has been shown to be the main lineage of Mesolithic central-west-north Europeans.
Ancient and modern mtDNA are great evidence that Mesolithic Europeans at least maternally largely descended of humans who arrived in Europe over 30,000 years ago. There are no Y DNA samples from Upper Palaeolithic Europe, but the fact that Y DNA I is most diverse in Europe, 6/7 Mesolithic Europeans tested have Y DNA I, and that it is estimated to be 20,000-25,000 years old is great evidence that many Upper Palaeolithic Europeans belonged to Y DNA I. There are other ancient and exclusively European Y DNA haplogroups like C1a2-V20 which is what Mesolithic Spaniard La Brana-1 had, but also F-96 and maybe others i don't know of. Upper Palaeolithic Siberian MA1 belonged to Y DNA haplogroup R* and he was a pure west Eurasian, it is possibly that some Upper Paleolithic European belonged to Y DNA R, Q, or another descendant of P.
Autosomal DNA shows that Mesolithic Europeans all descended from the same ancient source, that their modern descendants are largely confined to Europe, and that they are likely descended of very early Europeans who arrived i think at least before the last Ice age.
Not surprisingly modern Sardinians share much more dirft with Gok2 than any other modern populations. I was shocked to see that Sami(specifically from Norrbotten, Sweden) share much more drift with Ajv58 than any other modern Europeans, they share as much drift with Ajv58 as Sardinians do with Gok2. This suggests that Sami have the highest amount of Mesolithic European hunter gatherer ancestry today, and coincidentally are the last Europeans who are traditionally hunter gatherers.
Ajv58 may have had no farmer ancestry but Gok2 probably had around 30-40% hunter gatherer ancestry. So the D-statistics mean that Mesolithic European hunter gatherer ancestry is probably over 40% in much of northern and central Europe.
Skoglund et al. 2014 created admixture graphs like Lazaridis 2014 with ancient and modern samples, and came to the same conclusions.
Four key admixture events shown by their tree
Their admixture graphs like Lazardis suggest early European farmers harbored a significant amount of basal Eurasian ancestry(44+or-3% for Otzi, 23+or-6% for Gok2) and the rest of their ancestry formed a clad with European hunter gatherers. Most of their European hunter gatherer-like ancestry, descends from near easterns who were closely related to Mesolithic Europeans. Gok2 though is estimated to have 21% more than Otzi, and that 21% extra is defintley European hunter gatherer ancestry, Gok2 may have been as much as 40% Mesolithic European.
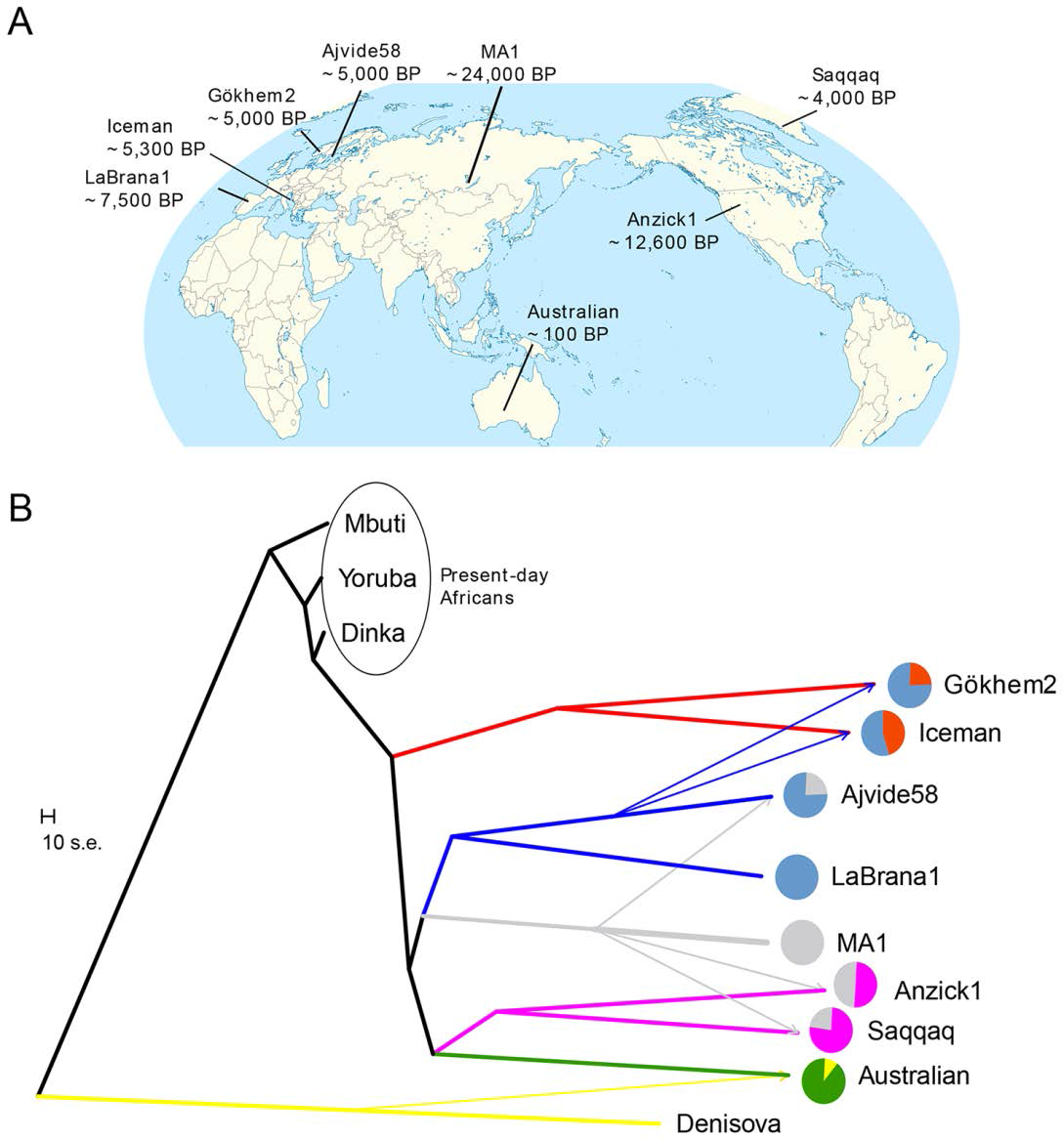
In their fitted model Ajv58(not La Brana-1) is the best fit for the hunter gatherers that contributed ancestry to Gok2 and Otzi. This is because Gok2's and Otzi's hunter gatherer ancestors were probably from central Europe(for Gok2 also northern Europe and Scandinavia) and were more related to Ajv58 a Swedish hunter gatherer than to La Brana-1 an Iberian hunter gatherer. Similarly Lazardis found that Stuttgart is more related to Loschbour and Motala12 than to La Brana-1, because her hunter gatherer ancestors were also probably from around central Europe.
Skoglund et al. 2014 fitted Ajv58 a Neolithic Swedish hunter gatherer as a mixture of MA1 and La Brana-1, like how Lazaridis 2014 fit Mesolithic Swedish hunter gatherer Motala12 as a mixture of MA1 and Loschbour. The predicted percentages of ANE(~25%) and WHG(~75%) ancestry in Ajv58 are very similar to what Lazaridis 2014 predicted for Motala12(19% ANE, 81% WHG). It was already noticed by Laz that Neolithic Swedish hunter gatherers cluster very closely to Mesolithic ones because they shift more towards MA1 in PCAs than La Brana-1 and Loschbour do. La Brana-1 like Loschbour did not fit as having any ANE ancestry.
This PCA of the ancient genomes with modern west Eurasians, are constant with Lazardis's and Davidski's(Eurogenes blog) PCAs.
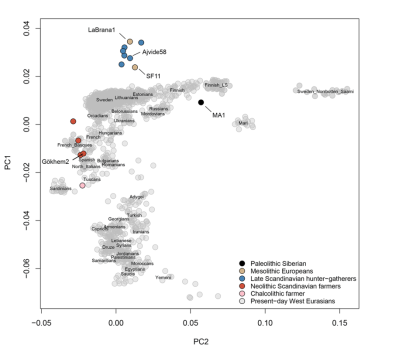
24,000BP Siberian MA1 clusters more closely with European hunter gatherers than with near easterns, which is probably because he and European hunter gatherers(some farmer ancestry for La Brana-1, and probably for the Neolithic Swedish hunter gatherers) are pure west Eurasians, unlike middle eastern specific ancestry which has a high amount of basal Eurasian.
The Neolithic Swedish farmers are much farther up towards European hunter gatherers than Otzi is. They cluster most closely with Basque, who are one of few modern Europeans who fitted as a mix of Stuttgart and Loschbour in Laz, and have the highest amount of WHG ancestry in southern Europe. One of the Neolithic Swedish farmers is shifted about as far up as central-north Europeans. In my opinion the Swedish Neolithic farmers had 30%- 40% WHG ancestry. They may have more hunter gatherer ancestry than Otzi and Stuttgart because they were the first farmers in Scandinavia.
The D-statistic above shows that MA1 is much closer to Lithuanians(arguable the most Mesolithic-like modern Europeans) than to Sardinians(the most Neolithic-like modern Europeans), like European hunter gatherers. He and StoraForvar11 are almost exactly as distant from Sardinians but La Brana-1 and the Swedish Neolithic hunter gatherers are less distant from Sardinians, probably because they had some farmer ancestry. MA1 has constantly been shown to be a close relative of stone age European hunter gatherers, but this is because he and stone age European hunter gatherers are the only samples known of pure or close to pure west Eurasians. Middle eastern-specific ancestry comes primarily from a source more related to European hunter gatherers than MA1 is, but its basal Eurasian ancestry makes them less related to European hunter gatherers than MA1 is.
Ajv58 had ancestral "dark skin" alleles in SNPs rs1426654, like La Brana-1 and Loschbour but unlike Mesolithic Swede Motala12 who had the derived "light skin" alleles. Ajv58 also had ancestral 'dark skin" alleles in SNP rs16891982 like Loschbour and La Brana-1 but unlike Mesolithic Swede Sf11 who had the derived "light skin" alleles. So most Mesolithic Europeans had the ancestral "dark skin" alleles in those two SNPs, but a minority had the derived "light skin" alleles.
Early European farmers Otzi, Gok2, and Stuttgart all had the derived "light skin" alleles in SNP rs1426654, Stuttgart had the ancestral 'dark skin" alleles in SNP rs16891982, while Gok2 and Otzi had the derived light skin alleles in SNP rs16891982. "Light skin" alleles in SNP rs1426654 were probably fixated in early European farmers, as they are in modern Europeans and west Asians. SNP rs16891982 is also associated with hair color, one study claims that if a European individual has the ancestral alleles there is a 7x better chance that individual has black hair. That's why the derived alleles are less popular in southern Europe than in northern Europe, and less popular in the middle east than in Europe.
Maybe since these two "light skin" mutations existed in both European hunter gatherers and farmers but at differnt rates, when the two populations mixed their descendants had these mutations at the same rate of their farmer ancestors. Like i have said in this thread and other times the science behind human skin color is not very well known and it's impossible to say what skin color these ancient people had.
I think i may have relatives who are good proxies for the skin color of most Mesolithic Europeans had. My brown skinned uncle is 92% European, 8% Native American+African, and he is missing the Ala111Thr and Phe374Leu mutations like La Brana-1, Loschbour, and Ajv58, and he is missing other mutations associated with European light skin and are fixated in modern Europeans. He is probably one of few people who have a significant amount of European hunter gatherer ancestry(probably around 40% or more) and is missing many of the same light skin mutations.
Neolithic European farmers defintley had light skin, since they had all the mutations associated with light skin in west Eurasia today and modern Sardinians are pracituclley no differnt genetically are generally light skinned. I tend towards saying stone European hunter gatherers had dark skin, because it cant be random that they are missing nearly every mutations associated with European light skin that are fixated in modern Europeans. There are alot of possibilities though, maybe both hunter gatherers and farmers had light skin. Today it seems all light pigmentation in Europe correlates with WHG ancestry, so possibly the hunter gatherers had light skin and some light hair, not just light eyes.
I was hoping Skoglund et al. 2014 would help find the origin of high amounts of light hair in modern Europeans. In supplementary pages 36-37 they have a horrible pigmentation predictor, so that's disappointing. In my opinion light hair in Europe today is either mainly descended of European hunter gatherers, or first became popular in hunter/farmer mutts. Since 3/4 Stone age European hunter gatherers tested so far are missing the L374F they were probably very dark haired. Near eastern ancestry today in Europe correlates with dark hair, Sardinians are the most early European farmer descended modern Europeans and are also the darkest haired Europeans so early European farmers were also probably very dark haired.
Mesolithic Swedish hunter gatherers
6,873 ± 119 BC, Stora Karlso Sweden : mtDNA=U4b1
Motala1(Female), 6,000BC Motala Sweden: mtDNA=U5a1
Motala2(Male), 6,000BC Motala Sweden: Y DNA=I* (I P38+, I PF3742+, I L41+, I1 S108-, I1 L845-, I1 M253-, I2a1b CT1293-, I2a2 L37-), mtDNA=U2e1
Motala3(Male) 6,000BC Motala Sweden: Y DNA=I2a1b*(I M258+, I PF3742+, I2 L68+, I2a1 P37.2+, I2a1b CTS7218+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b1 M359.2-, I2a1b3 L621-), mtDNA=U5a1
Motala4(Female) 6,000BC Motala Sweden: mtDNA=U5a2d
Motala6(Male) 6,000BC Motala Sweden: Y DNA=? (Q1 L232- Q1a2a L55+), mtDNA=U5a2d
Motala9(Male) 6,000BC Motala Sweden: Y DNA=I* (I P38+, I1 P40-), mtDNA=U5a2
Motala12(Male) 6,000BC Motala Sweden: Y DNA=pre-I2a1b or brother lineage to I2a1b(I PF3742+, I M258+, I M170+, I2 L68+, I2a L460+, I2a1 P37.2+, I2a1b CTS7218+, I2a1b CTS5985+. I2a1b L178+, I2a1b CTS1293+, I2a1b CTS176+, I2a1b CTS5375-, I2a1b CTS8486-, I2a1b1 M359.2-, I2a1b3 L621-), mtDNA=U2e1
Neolithic Swedish hunter gatherers of the Pitted Ware culture
Ajv52A(Male), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=V
Ajv59(Male), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U
Ajv53(Female), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4d
Ajv58(Male), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: Y DNA=I2a1-P37.2, mtDNA=U4d
Ajv70(Male), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4d
Ire8(Male), 5,100-4,150 cal. B.P, Ire, Hangvar, Gotland Sweden: mtDNA=U4d
Ajv13(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4
Ajv52b(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4
Ajv66(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U4
Ajv54(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U5
Ajv36(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U5
Ajv5(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U5a
Ajv29a(?), 4,900-4,600 cal B.P, Ajvide, Eksta, Gotland Sweden: mtDNA=U5a
Fir15(?), 2800-2000 BC,Fridtorp, Västerhejde, Gotland: mtDNA=U4
Fir22(?), 2800-2000 BC,Fridtorp, Västerhejde, Gotland: mtDNA=U4
Fir4(?), 2800-2000 BC,Fridtorp, Västerhejde, Gotland: mtDNA=U5
Fir27(?), 2800-2000 BC,Fridtorp, Västerhejde, Gotland: mtDNA=U5a
Ire6b(?), 5,100-4,150 cal. B.P, Ire, Hangvar, Gotland Sweden: mtDNA=T2b
Ire9(?), 5,100-4,150 cal. B.P, Ire, Hangvar, Gotland Sweden: mtDNA=U4
Ire3(?), 5,100-4,150 cal. B.P, Ire, Hangvar, Gotland Sweden: mtDNA=U4
Gökhem4(Male), 5,050-4,750 cal. years B.P.: mtDNA=H
Gökhem2(Female), 5,050-4,750 cal. years B.P.: mtDNA=H1c
Gökhem7(Female), 5,050-4,750 cal. years B.P.: mtDNA=H24
Gökhem5(Female), 5,280-4,890 cal. B.P.: mtDNA=K1e
Ste7(Female), 5,280-4,890 cal. B.P.: mtDNA=T2b
Ste7(Female), 5,280-4,890 cal. B.P.: mtDNA=J
The mtDNA of the Stone age Swedish farmers and hunter gatherers is 100% constant with what has been found in stone age farmers and hunter gatherers from other regions of Europe. The hunter gatherers have about 100% U5, U4, and U2, and the farmers mainly have modern European-specfic subclades of haplogroups which are most diverse and originated in the middle east. Since mDNA H1 and H3 are most popular in western Europe today, i bet that Neolithic west Europeans like the ones in Sweden had a very high amount and mtDNA samples from them so far is great evidence this is true. So far all U5 samples from Swedish hunter gatherers are U5a(mainly U5a1, but also U5a2 and U5a2d), like Mesolithic Russians(specifically U5a1) and unlike Mesolithic central-west Europeans who had mainly U5b(mainly U5b2) and the only U5a subclade found are U5a2(including U5a2c3* and U5a2a). The U5a in Swedish hunter gatherers may be from east European-very ANE like ancestors,, while their Y DNA I2a1-P37.2 is probably descended of central-west European ancestors.
At least paternally modern Balts and Scandinavians don't show evidence of a high amount of Mesolithic Swedish ancestry. I2a1-P37.2 today is very very rare in Scandnavians and Balts(much of it is probably east European=specfic I2a1b1a-L147.2), there are some Scandinavian P37.2 samples in FTDNA's database who have not been tested for I2a1-P37.2 subclades i bet at least have unknown specific Scandinavian SNPs and are a brotherclade to I2a1b. Y DNA I2a1-P37.2 is likely a marker of Europeans who took refuge in south-western Europe during the last ice age, today it is most diverse in western Europe and so far has been shown to be the main lineage of Mesolithic central-west-north Europeans.
Ancient and modern mtDNA are great evidence that Mesolithic Europeans at least maternally largely descended of humans who arrived in Europe over 30,000 years ago. There are no Y DNA samples from Upper Palaeolithic Europe, but the fact that Y DNA I is most diverse in Europe, 6/7 Mesolithic Europeans tested have Y DNA I, and that it is estimated to be 20,000-25,000 years old is great evidence that many Upper Palaeolithic Europeans belonged to Y DNA I. There are other ancient and exclusively European Y DNA haplogroups like C1a2-V20 which is what Mesolithic Spaniard La Brana-1 had, but also F-96 and maybe others i don't know of. Upper Palaeolithic Siberian MA1 belonged to Y DNA haplogroup R* and he was a pure west Eurasian, it is possibly that some Upper Paleolithic European belonged to Y DNA R, Q, or another descendant of P.
Autosomal DNA shows that Mesolithic Europeans all descended from the same ancient source, that their modern descendants are largely confined to Europe, and that they are likely descended of very early Europeans who arrived i think at least before the last Ice age.
Autosomal DNA
By using D-statistics Skoglund et al. 2014 found that Ajv58 fits as a clade with La brana-1, modern Sardinians fit as a clade with Otzi, and Gok2 has more hunter gatherer ancestry than Otzi. They found that all non-west Eurasian non Africans share more drift with Ajv58, which is because Gok2 had some basal Eurasian ancestry which does not have the shared drift between west and east Eurasians. The modern Europeans who share more dirft with either Ajv58 or Gok2 is constant with how Mesolithic European and near eastern ancestry is distributed in Europe. Northern and central Europeans are closer to Ajv58 while southern Europeans(incl. French) are closer to Gok2. Not surprisingly modern Sardinians share much more dirft with Gok2 than any other modern populations. I was shocked to see that Sami(specifically from Norrbotten, Sweden) share much more drift with Ajv58 than any other modern Europeans, they share as much drift with Ajv58 as Sardinians do with Gok2. This suggests that Sami have the highest amount of Mesolithic European hunter gatherer ancestry today, and coincidentally are the last Europeans who are traditionally hunter gatherers.
Ajv58 may have had no farmer ancestry but Gok2 probably had around 30-40% hunter gatherer ancestry. So the D-statistics mean that Mesolithic European hunter gatherer ancestry is probably over 40% in much of northern and central Europe.
Skoglund et al. 2014 created admixture graphs like Lazaridis 2014 with ancient and modern samples, and came to the same conclusions.
Four key admixture events shown by their tree
1. Gene flow from Denisovans to Australian aboriginal populations (48, 97, 98)
2. Gene flow from the MA1 lineage to Native American ancestors (Anzick1 and
Saqqaq) (13)
3. Gene flow from the MA1 lineage to Ajvide58, see main text and Table S13.
4. Gene flow from Late Mesolithic hunter-gatherers (La Brana 1) to early
farmers (Gökhem2 and Iceman) (18).
Their admixture graphs like Lazardis suggest early European farmers harbored a significant amount of basal Eurasian ancestry(44+or-3% for Otzi, 23+or-6% for Gok2) and the rest of their ancestry formed a clad with European hunter gatherers. Most of their European hunter gatherer-like ancestry, descends from near easterns who were closely related to Mesolithic Europeans. Gok2 though is estimated to have 21% more than Otzi, and that 21% extra is defintley European hunter gatherer ancestry, Gok2 may have been as much as 40% Mesolithic European.
In their fitted model Ajv58(not La Brana-1) is the best fit for the hunter gatherers that contributed ancestry to Gok2 and Otzi. This is because Gok2's and Otzi's hunter gatherer ancestors were probably from central Europe(for Gok2 also northern Europe and Scandinavia) and were more related to Ajv58 a Swedish hunter gatherer than to La Brana-1 an Iberian hunter gatherer. Similarly Lazardis found that Stuttgart is more related to Loschbour and Motala12 than to La Brana-1, because her hunter gatherer ancestors were also probably from around central Europe.
Skoglund et al. 2014 fitted Ajv58 a Neolithic Swedish hunter gatherer as a mixture of MA1 and La Brana-1, like how Lazaridis 2014 fit Mesolithic Swedish hunter gatherer Motala12 as a mixture of MA1 and Loschbour. The predicted percentages of ANE(~25%) and WHG(~75%) ancestry in Ajv58 are very similar to what Lazaridis 2014 predicted for Motala12(19% ANE, 81% WHG). It was already noticed by Laz that Neolithic Swedish hunter gatherers cluster very closely to Mesolithic ones because they shift more towards MA1 in PCAs than La Brana-1 and Loschbour do. La Brana-1 like Loschbour did not fit as having any ANE ancestry.
This PCA of the ancient genomes with modern west Eurasians, are constant with Lazardis's and Davidski's(Eurogenes blog) PCAs.
24,000BP Siberian MA1 clusters more closely with European hunter gatherers than with near easterns, which is probably because he and European hunter gatherers(some farmer ancestry for La Brana-1, and probably for the Neolithic Swedish hunter gatherers) are pure west Eurasians, unlike middle eastern specific ancestry which has a high amount of basal Eurasian.
The Neolithic Swedish farmers are much farther up towards European hunter gatherers than Otzi is. They cluster most closely with Basque, who are one of few modern Europeans who fitted as a mix of Stuttgart and Loschbour in Laz, and have the highest amount of WHG ancestry in southern Europe. One of the Neolithic Swedish farmers is shifted about as far up as central-north Europeans. In my opinion the Swedish Neolithic farmers had 30%- 40% WHG ancestry. They may have more hunter gatherer ancestry than Otzi and Stuttgart because they were the first farmers in Scandinavia.
The D-statistic above shows that MA1 is much closer to Lithuanians(arguable the most Mesolithic-like modern Europeans) than to Sardinians(the most Neolithic-like modern Europeans), like European hunter gatherers. He and StoraForvar11 are almost exactly as distant from Sardinians but La Brana-1 and the Swedish Neolithic hunter gatherers are less distant from Sardinians, probably because they had some farmer ancestry. MA1 has constantly been shown to be a close relative of stone age European hunter gatherers, but this is because he and stone age European hunter gatherers are the only samples known of pure or close to pure west Eurasians. Middle eastern-specific ancestry comes primarily from a source more related to European hunter gatherers than MA1 is, but its basal Eurasian ancestry makes them less related to European hunter gatherers than MA1 is.
Pigmentation
Ajv58 had ancestral "dark skin" alleles in SNPs rs1426654, like La Brana-1 and Loschbour but unlike Mesolithic Swede Motala12 who had the derived "light skin" alleles. Ajv58 also had ancestral 'dark skin" alleles in SNP rs16891982 like Loschbour and La Brana-1 but unlike Mesolithic Swede Sf11 who had the derived "light skin" alleles. So most Mesolithic Europeans had the ancestral "dark skin" alleles in those two SNPs, but a minority had the derived "light skin" alleles.
Early European farmers Otzi, Gok2, and Stuttgart all had the derived "light skin" alleles in SNP rs1426654, Stuttgart had the ancestral 'dark skin" alleles in SNP rs16891982, while Gok2 and Otzi had the derived light skin alleles in SNP rs16891982. "Light skin" alleles in SNP rs1426654 were probably fixated in early European farmers, as they are in modern Europeans and west Asians. SNP rs16891982 is also associated with hair color, one study claims that if a European individual has the ancestral alleles there is a 7x better chance that individual has black hair. That's why the derived alleles are less popular in southern Europe than in northern Europe, and less popular in the middle east than in Europe.
Maybe since these two "light skin" mutations existed in both European hunter gatherers and farmers but at differnt rates, when the two populations mixed their descendants had these mutations at the same rate of their farmer ancestors. Like i have said in this thread and other times the science behind human skin color is not very well known and it's impossible to say what skin color these ancient people had.
I think i may have relatives who are good proxies for the skin color of most Mesolithic Europeans had. My brown skinned uncle is 92% European, 8% Native American+African, and he is missing the Ala111Thr and Phe374Leu mutations like La Brana-1, Loschbour, and Ajv58, and he is missing other mutations associated with European light skin and are fixated in modern Europeans. He is probably one of few people who have a significant amount of European hunter gatherer ancestry(probably around 40% or more) and is missing many of the same light skin mutations.
Neolithic European farmers defintley had light skin, since they had all the mutations associated with light skin in west Eurasia today and modern Sardinians are pracituclley no differnt genetically are generally light skinned. I tend towards saying stone European hunter gatherers had dark skin, because it cant be random that they are missing nearly every mutations associated with European light skin that are fixated in modern Europeans. There are alot of possibilities though, maybe both hunter gatherers and farmers had light skin. Today it seems all light pigmentation in Europe correlates with WHG ancestry, so possibly the hunter gatherers had light skin and some light hair, not just light eyes.
I was hoping Skoglund et al. 2014 would help find the origin of high amounts of light hair in modern Europeans. In supplementary pages 36-37 they have a horrible pigmentation predictor, so that's disappointing. In my opinion light hair in Europe today is either mainly descended of European hunter gatherers, or first became popular in hunter/farmer mutts. Since 3/4 Stone age European hunter gatherers tested so far are missing the L374F they were probably very dark haired. Near eastern ancestry today in Europe correlates with dark hair, Sardinians are the most early European farmer descended modern Europeans and are also the darkest haired Europeans so early European farmers were also probably very dark haired.
Last edited: