Geographic distribution
Distribution of haplogroup R1b in Europe

R1b is the most common haplogroup in Western Europe, reaching over 80% of the population in Ireland, the Scottish Highlands, western Wales, the Atlantic fringe of France, the Basque country and Catalonia. It is also common in Anatolia and around the Caucasus, in parts of Russia and in Central and South Asia. Besides the Atlantic and North Sea coast of Europe, hotspots include the Po valley in north-central Italy (over 70%), Armenia (35%), the Bashkirs of the Urals region of Russia (50%), Turkmenistan (over 35%), the Hazara people of Afghanistan (35%), the Uyghurs of North-West China (20%) and the Newars of Nepal (11%). R1b-V88, a subclade specific to sub-Saharan Africa, is found in 60 to 95% of men in northern Cameroon.
Distribution map of haplogroup R1b in the Old World
 - Eupedia")
Phylogeny of R1b
If you are new to genetic genealogy, please check our Introduction to phylogenetics to understand how to read a phylogenetic tree.
 - Eupedia")
Click on the trees below to enlarge
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
Origins & History
Paleolithic mammoth hunters
Haplogroup R* originated in North Asia just before the Last Glacial Maximum (26,500-19,000 years ago). This haplogroup has been identified in the remains of a 24,000 year-old boy from the Altai region, in south-central Siberia (Raghavan et al. 2013). This individual belonged to a tribe of mammoth hunters that may have roamed across Siberia and parts of Europe during the Paleolithic. Autosomally this Paleolithic population appears to have contributed mostly to the ancestry of modern Europeans and South Asians, the two regions where haplogroup R also happens to be the most common nowadays (R1b in Western Europe, R1a in Eastern Europe, Central and South Asia, and R2 in South Asia).
The oldest forms of R1b (M343, P25, L389) are found dispersed at very low frequencies from Western Europe to India, a vast region where could have roamed the nomadic R1b hunter-gatherers during the Ice Age. The three main branches of R1b1 (R1b1a, R1b1b, R1b1c) all seem to have stemmed from the Middle East. The southern branch, R1b1c (V88), is found mostly in the Levant and Africa. The northern branch, R1b1a (P297), seems to have originated around the Caucasus, eastern Anatolia or northern Mesopotamia, then to have crossed over the Caucasus, from where they would have invaded Europe and Central Asia. R1b1b (M335) has only been found in Anatolia.
Neolithic cattle herders
It has been hypothetised that R1b people (perhaps alongside neighbouring J2 tribes) were the first to domesticate cattle in northern Mesopotamia some 10,500 years ago. R1b tribes descended from mammoth hunters, and when mammoths went extinct, they started hunting other large game such as bisons and aurochs. With the increase of the human population in the Fertile Crescent from the beginning of the Neolithic (starting 12,000 years ago), selective hunting and culling of herds started replacing indiscriminate killing of wild animals. The increased involvement of humans in the life of aurochs, wild boars and goats led to their progressive taming. Cattle herders probably maintained a nomadic or semi-nomadic existence, while other people in the Fertile Crescent (presumably represented by haplogroups E1b1b, G and T) settled down to cultivate the land or keep smaller domesticates.
The analysis of bovine DNA has revealed that all the taurine cattle (Bos taurus) alive today descend from a population of only 80 aurochs. The earliest evidence of cattle domestication dates from circa 8,500 BCE in the Pre-Pottery Neolithic cultures in the Taurus Mountains. The two oldest archaeological sites showing signs of cattle domestication are the villages of Çayönü Tepesi in southeastern Turkey and Dja'de el-Mughara in northern Iraq, two sites only 250 km away from each others. This is presumably the area from which R1b lineages started expanding - or in other words the "original homeland" of R1b.
The early R1b cattle herders would have split in at least three groups. One branch (M335) remained in Anatolia, but judging from its extreme rarity today wasn't very successful, perhaps due to the heavy competition with other Neolithic populations in Anatolia, or to the scarcity of pastures in this mountainous environment. A second branch migrated south to the Levant, where it became the V88 branch. Some of them searched for new lands south in Africa, first in Egypt, then colonising most of northern Africa, from the Mediterranean coast to the Sahel. The third branch (P297), crossed the Caucasus into the vast Pontic-Caspian Steppe, which provided ideal grazing grounds for cattle. They split into two factions: R1b1a1 (M73), which went east along the Caspian Sea to Central Asia, and R1b1a2 (M269), which at first remained in the North Caucasus and the Pontic Steppe between the Dnieper and the Volga. It is not yet clear whether M73 actually migrated across the Caucasus and reached Central Asia via Kazakhstan, or if it went south through Iran and Turkmenistan. In any case, M73 would be a pre-Indo-European branch of R1b, just like V88 and M335.
R1b-M269 (the most common form in Europe) is closely associated with the diffusion of Indo-European languages, as attested by its presence in all regions of the world where Indo-European languages were spoken in ancient times, from the Atlantic coast of Europe to the Indian subcontinent, which comprised almost all Europe (except Finland, Sardinia and Bosnia-Herzegovina), Anatolia, Armenia, European Russia, southern Siberia, many pockets around Central Asia (notably in Xinjiang, Turkmenistan, Tajikistan and Afghanistan), without forgetting Iran, Pakistan, northern India and Nepal. The history of R1b and R1a are intricately connected to each others.
The Levantine & African branch of R1b (V88)
Like its northern counterpart (R1b-M269), R1b-V88 is associated with the domestication of cattle in northern Mesopotamia. Both branches of R1b probably split soon after cattle were domesticated, approximately 10,500 years ago (8,500 BCE). R1b-V88 migrated south towards the Levant and Egypt. The migration of R1b people can be followed archeologically through the presence of domesticated cattle, which appear in central Syria around 8,000-7,500 BCE (late Mureybet period), then in the Southern Levant and Egypt around 7,000-6,500 BCE (e.g. at Nabta Playa and Bir Kiseiba). Cattle herders subsequently spread across most of northern and eastern Africa. The Sahara desert would have been more humid during the Neolithic Subpluvial period (c. 7250-3250 BCE), and would have been a vast savannah full of grass, an ideal environment for cattle herding.
Evidence of cow herding during the Neolithic has shown up at Uan Muhuggiag in central Libya around 5500 BCE, at the Capeletti Cave in northern Algeria around 4500 BCE. But the most compelling evidence that R1b people related to modern Europeans once roamed the Sahara is to be found at Tassili n'Ajjer in southern Algeria, a site famous pyroglyphs (rock art) dating from the Neolithic era. Some painting dating from around 3000 BCE depict fair-skinned and blond or auburn haired women riding on cows. The oldest known R1b-V88 sample in Europe is a 6,200 year-old farmer/herder from Catalonia tested by Haak et al. (2015). Autosomally this individual was a typical Near Eastern farmer, possessing just a little bit of Mesolithic West European admixture.
After reaching the Maghreb, R1b-V88 cattle herders could have crossed the Strait of Gibraltar to Iberia, probably accompanied by G2 farmers, J1 and T1a goat herders. These North African Neolithic farmers/herders could have been the ones who established the Almagra Pottery culture in Andalusia in the 6th millennium BCE.
Nowadays small percentages (1 to 4%) of R1b-V88 are found in the Levant, among the Lebanese, the Druze, and the Jews, and almost in every country in Africa north of the equator. Higher frequency in Egypt (5%), among Berbers from the Egypt-Libya border (23%), among the Sudanese Copts (15%), the Hausa people of Sudan (40%), the the Fulani people of the Sahel (54% in Niger and Cameroon), and Chadic tribes of northern Nigeria and northern Cameroon (especially among the Kirdi), where it is observed at a frequency ranging from 30% to 95% of men. According to Cruciani et al. (2010) R1b-V88 would have crossed the Sahara between 9,200 and 5,600 years ago, and is most probably associated with the diffusion of Chadic languages, a branch of the Afroasiatic languages. V88 would have migrated from Egypt to Sudan, then expanded along the Sahel until northern Cameroon and Nigeria. However, R1b-V88 is not only present among Chadic speakers, but also among Senegambian speakers (Fula-Hausa) and Semitic speakers (Berbers, Arabs).
R1b-V88 is found among the native populations of Rwanda, South Africa, Namibia, Angola, Congo, Gabon, Equatorial Guinea, Ivory Coast, Guinea-Bissau. The wide distribution of V88 in all parts of Africa, its incidence among herding tribes, and the coalescence age of the haplogroup all support a Neolithic dispersal. In any case, a later migration out of Egypt would be improbable since it would have brought haplogroups that came to Egypt during the Bronze Age, such as J1, J2, R1a or R1b-L23.
The maternal lineages associated with the spread of R1b-V88 in Africa are mtDNA haplogroups J1b, U5 and V, and perhaps also U3 and some H subclades (=> see Retracing the mtDNA haplogroups of the original R1b people).
The North Caucasus and the Pontic-Caspian steppe : the Indo-European link
Modern linguists have placed the Proto-Indo-European homeland in the Pontic-Caspian Steppe, a distinct geographic and archeological region extending from the Danube estuary to the Ural mountains to the east and North Caucasus to the south. The Neolithic, Eneolithic and early Bronze Age cultures in Pontic-Caspian steppe has been called the Kurgan culture (4200-2200 BCE) by Marija Gimbutas, due to the lasting practice of burying the deads under mounds ("kurgan") among the succession of cultures in that region. It is now known that kurgan-type burials only date from the 4th millenium BCE and almost certainly originated south of the Caucasus. The genetic diversity of R1b being greater around eastern Anatolia, it is hard to deny that R1b evolved there before entering the steppe world.
Horses were first domesticated around 4600 BCE in the Caspian Steppe, perhaps somewhere around the Don or the lower Volga, and soon became a defining element of steppe culture. Nevertheless it is unlikely that R1b was already present in the eastern steppes at the time, so the domestication of the horse should be attributed to the indigenous R1a people, or tribes belonging to the older R1b-P297 branch, which settled in eastern Europe during the Late Paleolithic or Mesolithic period. Samples from Mesolithic Samara (Haak 2015) and Latvia (Jones 2017) all belonged to R1b-P297. Autosomally these Mesolithic R1a and R1b individuals were nearly pure Mesolithic East European, sometimes with a bit of Siberian admixture, but lacked the additional Caucasian admixture found in the Chalcolithic Afanasevo, Yamna and Corded Ware samples.
It is not yet entirely clear when R1b-M269 crossed over from the South Caucasus to the Pontic-Caspian steppe. This might have happened with the appearance of the Dnieper-Donets culture (c. 5100-4300 BCE). This was the first truly Neolithic society in the Pontic-Caspian Steppe. Domesticated animals (cattle, sheep and goats) were herded throughout the steppes and funeral rituals were elaborate. Sheep wool would play an important role in Indo-European society, notably in the Celtic and Germanic (R1b branches of the Indo-Europeans) clothing traditions up to this day. However, many elements indicate a continuity in the Dnieper-Donets culture with the previous Mesolithic hunter-gatherers, and at the same time an influence from the Balkans and Carpathians, with regular imports of pottery and copper objects. It is therefore more likely that Dnieper-Donets marked the transition of indigenous R1a and/or I2a1b people to early agriculture, perhaps with an influx of Near Eastern farmers from 'Old Europe'. Over 30 DNA samples from Neolithic Ukraine (5500-4800 BCE) were tested by Mathieson et al. (2017). They belonged to Y-haplogroups I, I2a2, R1a, R1b1a (L754) and one R1b1a2 (L388). None of them belonged to R1b-M269 or R1b-L23 clades, which dominated during the Yamna period. Mitochondrial lineages were also exclusively of Mesolithic European origin (U4a, U4b, U4d, U5a1, U5a2, U5b2, as well as one J2b1 and one U2e1). None of those maternal lineages include typical Indo-European haplogroups, like H2a1, H6, H8, H15, I1a1, J1b1a, W3, W4 or W5 that would later show up in the Yamna, Corded Ware and Unetice cultures. Indeed, autosomally genomes from Neolithic Ukraine were purely Mesolithic European (about 90% EHG and 10% WHG) and completely lacked the Caucasian (CHG) admxiture later found in Yamna and subsequent Indo-European cultures during the Bronze Age.
The first clearly Proto-Indo-European cultures were the Khvalynsk (5200-4500 BCE) and Sredny Stog (4600-3900 BCE) cultures in the Pontic-Caspian Steppe. This is when small kurgan burials begin to appear, with the distinctive posturing of the dead on the back with knees raised and oriented toward the northeast, which would be found in later steppe cultures as well. There is evidence of population blending from the variety of skull shapes. Towards the end of the 5th millennium, an elite starts to develop with cattle, horses and copper used as status symbols. It is at the turn of the Khvalynsk and Sredny Stog periods that R1b-M269's main subclade, L23, is thought to have appeared, around 4,500 BCE. 99% of Indo-European R1b descends from this L23 clade. The other branch descended from M269 is PF7562, which is found mostly in the Balkans, Turkey and Armenia today, and may represent an early Steppe migration to the Balkans dating from the Sredny Stog period.
Another migration across the Caucasus happened shortly before 3700 BCE, when the Maykop culture, the world's first Bronze Age society, suddenly materialised in the north-west Caucasus, apparently out of nowhere. The origins of Maykop are still uncertain, but archeologists have linked it to contemporary Chalcolithic cultures in Assyria and western Iran. Archeology also shows a clear diffusion of bronze working and kurgan-type burials from the Maykop culture to the Pontic Steppe, where the Yamna culture developed soon afterwards (from 3500 BCE). Kurgan (a.k.a. tumulus) burials would become a dominant feature of ancient Indo-European societies and were widely used by the Celts, Romans, Germanic tribes, and Scythians, among others.
The Yamna period (3500-2500 BCE) is the most important one in the creation of Indo-European culture and society. Middle Eastern R1b-M269 people had been living and blending to some extent with the local R1a foragers and herders for over a millennium, perhaps even two or three. The close cultural contact and interactions between R1a and R1b people all over the Pontic-Caspian Steppe resulted in the creation of a common vernacular, a new lingua franca, which linguists have called Proto-Indo-European (PIE). It is pointless to try to assign another region of origin to the PIE language. Linguistic similarities exist between PIE and Caucasian and Hurrian languages in the Middle East on the one hand, and Uralic languages in the Volga-Ural region on the other hand, which makes the Pontic Steppe the perfect intermediary region.
During the Yamna period cattle and sheep herders adopted wagons to transport their food and tents, which allowed them to move deeper into the steppe, giving rise to a new mobile lifestyle that would eventually lead to the great Indo-European migrations. This type of mass migration in which whole tribes moved with the help of wagons was still common in Gaul at the time of Julius Caesar, and among Germanic peoples in the late Antiquity.
The Yamna horizon was not a single, unified culture. In the south, along the northern shores of the Black Sea coast until the the north-west Caucasus, was a region of open steppe, expanding eastward until the Caspian Sea, Siberia and Mongolia (the Eurasian Steppe). The western section, between the Don and Dniester Rivers (and later the Danube), was the one most densely settled by R1b people, with only a minority of R1a people (5-10%). The eastern section, in the Volga basin until the Ural mountains, was inhabited by R1a people with a substantial minority of R1b people (whose descendants can be found among the Bashkirs, Turkmans, Uyghurs and Hazaras, among others). The northern part of the Yamna horizon was forest-steppe occupied by R1a people, also joined by a small minority of R1b (judging from Corded Ware samples and from modern Russians and Belarussians, whose frequency of R1b is from seven to nine times lower than R1a). The western branch would migrate to the Balkans and Greece, then to Central and Western Europe, and back to their ancestral Anatolia in successive waves (Hittites, Phrygians, Armenians, etc.). The eastern branch would migrate to Central Asia, Xinjiang, Siberia, and South Asia (Iran, Pakistan, India). The northern branch would evolve into the Corded Ware culture and disperse around the Baltic, Poland, Germany and Scandinavia.
The Maykop culture, the R1b link to the Steppe?
The Maykop culture (3700-2500 BCE) in the north-west Caucasus was culturally speaking a sort of southern extension of the Yamna horizon. Although not generally considered part of the Pontic-Caspian steppe culture due to its geography, the North Caucasus had close links with the steppes, as attested by numerous ceramics, gold, copper and bronze weapons and jewelry in the contemporaneous cultures of Mikhaylovka, Sredny Stog and Kemi Oba. The link between the northern Black Sea coast and the North Caucasus is older than the Maykop period. Its predecessor, the Svobodnoe culture (4400-3700 BCE), already had links to the Suvorovo-Novodanilovka and early Sredny Stog cultures. The even older Nalchik settlement (5000-4500 BCE) in the North Caucasus displayed a similar culture as Khvalynsk in the Caspian Steppe and Volga region. This may be the period when R1b started interracting and blending with the R1a population of the steppes.
The Yamna and Maykop people both used kurgan burials, placing their deads in a supine position with raised knees and oriented in a north-east/south-west axis. Graves were sprinkled with red ochre on the floor, and sacrificed domestic animal buried alongside humans. They also had in common horses, wagons, a heavily cattle-based economy with a minority of sheep kept for their wool, use of copper/bronze battle-axes (both hammer-axes and sleeved axes) and tanged daggers. In fact, the oldest wagons and bronze artefacts are found in the North Caucasus, and appear to have spread from there to the steppes.
Maykop was an advanced Bronze Age culture, actually one of the very first to develop metalworking, and therefore metal weapons. The world's oldest sword was found at a late Maykop grave in Klady kurgan 31. Its style is reminiscent of the long Celtic swords, though less elaborated. Horse bones and depictions of horses already appear in early Maykop graves, suggesting that the Maykop culture might have been founded by steppe people or by people who had close link with them. However, the presence of cultural elements radically different from the steppe culture in some sites could mean that Maykop had a hybrid population. Without DNA testing it is impossible to say if these two populations were an Anatolian R1b group and a G2a Caucasian group, or whether R1a people had settled there too. The two or three ethnicities might even have cohabited side by side in different settlements. The one typical Caucasian Y-DNA lineage that does follow the pattern of Indo-European migrations is G2a-L13, which is found throughout Europe, Central Asia and South Asia. In the Balkans, the Danube basin and Central Europe its frequency is somewhat proportional to the percentage of R1b.
Maykop people are the ones credited for the introduction of primitive wheeled vehicles (wagons) from Mesopotamia to the Steppe. This would revolutionise the way of life in the steppe, and would later lead to the development of (horse-drawn) war chariots around 2000 BCE. Cavalry and chariots played an vital role in the subsequent Indo-European migrations, allowing them to move quickly and defeat easily anybody they encountered. Combined with advanced bronze weapons and their sea-based culture, the western branch (R1b) of the Indo-Europeans from the Black Sea shores are excellent candidates for being the mysterious Sea Peoples, who raided the eastern shores of the Mediterranean during the second millennium BCE.
The rise of the IE-speaking Hittites in Central Anatolia happened a few centuries after the disappearance of the Maykop and Yamna cultures. Considering that most Indo-European forms of R1b found in Anatolia today belong to the R1b-Z2103 subclade, it makes little doubt that the Hittites came to Anatolia via the Balkans, after Yamna/Maykop people invaded Southeast Europe. The Maykop and Yamna cultures were succeeded by the Srubna culture (1600-1200 BCE), possibly representing an advance of R1a-Z282 people from the northern steppes towards the Black Sea shores, filling the vacuum left by the R1b tribes who migrated to Southeast Europe and Anatolia.
Migration map of Y-haplogroup R1b from the Paleolithic to the end of the Bronze Age (c. 1000 BCE)

Migration map of Y-haplogroup R1b in the Late Neolithic and Early Bronze Age (c. 5500-3000 BCE)
The Siberian & Central Asian branch
When R1b crossed the Caucasus in the Late Neolithic, it split into two main groups. The western one (L51) would settle the eastern and northern of the Black Sea. The eastern one (Z2103) migrated to the Don-Volga region, where horses were domesticated circa 4600 BCE. R1b probably mixed with indigenous R1a people and founded the Repin culture (3700-3300 BCE) a bit before the Yamna culture came into existence in the western Pontic Steppe. R1b would then have migrated with horses along the Great Eurasian Steppe until the Altai mountains in East-Central Asia, where they established the Afanasevo culture (c. 3600-2400 BCE). Afanasevo people might be the precursors of the Tocharian branch of Indo-European languages. In 2014, Clément Hollard of Strasbourg University tested three Y-DNA samples from the Afanasevo culture and all three turned out to belong to haplogroup R1b, including two to R1b-M269.
The R1b people who stayed in the Volga-Ural region were probably the initiators of the Poltavka culture (2700-2100 BCE), then became integrated into the R1a-dominant Sintashta-Petrovka culture (2100-1750 BCE) linked to the Indo-Aryan conquest of Central and South Asia (=> see R1a for more details).
Nowadays in Russia R1b is found at higher frequencies among ethnic minorities of the Volga-Ural region (Udmurts, Komi, Mordvins, Tatars) than among Slavic Russians. R1b is also present in many Central Asian populations, the highest percentages being observed among the Uyghurs (20%) of Xinjiang in north-west China, the Yaghnobi people of Tajikistan (32%), and the Bashkirs (47%, or 62.5% in the Abzelilovsky district) of Bashkortostan in Russia (border of Kazakhstan).
R1b-M73, found primarily in North Asia (Altai, Mongolia), Central Asia and the North Caucasus is thought to have spread during the Neolithic from the Middle East to Central and North Asia, and therefore can be considered to be pre-Indo-European.
The European & Middle Eastern branch
The Indo-Europeans's bronze weapons and the extra mobility provided by horses would have given them a tremendous advantage over the autochthonous inhabitants of Europe, namely the native haplogroup C1a2, F and I (descendants of Cro-Magnon) and the early Neolithic herders and farmers (G2a, H2, E1b1b and T1a). This allowed R1a and R1b to replace most of the native male lineages (=> see How did R1b come to replace most of the older lineages in Western Europe?), although female lineages seem to have been less affected.
A comparison with the Indo-Iranian invasion of South Asia shows that 40% of the male linages of northern India are R1a, but less than 10% of the female lineages could be of Indo-European origin. The impact of the Indo-Europeans was more severe in Europe because European society 4,000 years ago was less developed in terms of agriculture, technology (no bronze weapons) and population density than that of the Indus Valley civilization. This is particularly true of the native Western European cultures where farming arrived much later than in the Balkans or Central Europe. Greece, the Balkans and the Carpathians were the most advanced of European societies at the time and were the least affected in terms of haplogroup replacement. neolithic lineages survived better in regions that were more difficult to reach or less hospitable to horse breeders, like the Alps, the Dinaric Alps, the Apennines and Sardinia.
The Conquest of "Old Europe" and Central Europe (4200-2500 BCE)
The first forays of Steppe people into the Balkans happened between 4200 BCE and 3900 BCE, when cattle herders equipped with horse-drawn wagons crossed the Dniester and Danube and apparently destroyed the towns of the Gumelnița, Varna and Karanovo VI cultures in Eastern Romania and Bulgaria. A climatic change resulting in colder winters during this exact period probably pushed steppe herders to seek milder pastures for their stock, while failed crops would have led to famine and internal disturbance within the Danubian and Balkanic communities. The ensuing Cernavodă culture (Copper Age, 4000-3200 BCE), Coțofeni/Usatovo culture (Copper to Bronze Age, 3500-2500 BCE), Ezero culture (Bronze Age, 3300-2700 BCE), in modern Romania, seems to have had a mixed population of steppe immigrants and people from the old tell settlements. These Steppe immigrants were likely a mixture of both R1a and R1b lineages, with a probably higher percentage of R1a than later Yamna-era invasions.
The Steppe invaders would have forced many Danubian farmers to migrate to the Cucuteni-Trypillian towns in the eastern Carpathians, causing a population boom and a north-eastward expansion until the Dnieper valley, bringing Y-haplogroups G2a, I2a1 (probably the dominant lineage of the Cucuteni-Trypillian culture), E1b1b, J2a and T1a in what is now central Ukraine. This precocious Indo-European advance westward was fairly limited, due to the absence of Bronze weapons and organised army at the time, and was indeed only possible thanks to climatic catastrophes which reduced the defences of the towns of Old Europe. The Carphatian, Danubian, and Balkanic cultures were too densely populated and technologically advanced to allow for a massive migration.
In comparison the forest-steppe R1a people successfully penetrated into the heart of Europe with little hindrance, due to the absence of developed agrarian societies around Poland and the Baltic. The Corded Ware culture (3200-1800 BCE) was a natural northern and western expansion of the Yamna culture, reaching as far west as Germany and as far north as Sweden and Norway. DNA analysis from the Corded Ware confirmed the presence of R1a and R1b in Poland c. 2700 BCE and R1a central Germany around 2600 BCE. The Corded Ware tribes expanded from the northern fringe of the Yamna culture where R1a lineages were prevalent over R1b ones.
The expansion of R1b people into Old Europe was slower, but proved inevitable. In 2800 BCE, by the time the Corded Ware had already reached Scandinavia, the Bronze Age R1b cultures had barely moved into the Pannonian Steppe. They established major settlements in the Great Hungarian Plain, the most similar habitat to their ancestral Pontic Steppes. Around 2500 BCE, the western branch of Indo-European R1b were poised for their next major expansion into modern Germany and Western Europe. By that time, the R1b immigrants had blended to a great extent with the indigenous Mesolithic and Neolithic populations of the Danubian basin, where they had now lived for 1,700 years.
The strongly partriarchal Indo-European elite remained almost exclusively R1b on the paternal side, but absorbed a high proportion of non-Indo-European maternal lineages. Hybridised, the new Proto-Indo-European R1b people would have lost most of their remaining Proto-Europoid or Mongolid features inherited from their Caspian origins (which were still clearly visible in numerous individuals from the Yamna period). Their light hair, eye and skin pigmentation, once interbred with the darker inhabitants of Old Europe, became more like that of modern Southern Europeans. The R1a people of the Corded Ware culture would come across far less populous societies in Northern Europe, mostly descended from the lighter Mesolithic population, and therefore retained more of their original pigmentation (although facial traits evolved considerably in Scandinavia, where the I1 inhabitants were strongly dolicocephalic and long-faced, as opposed to the brachycephalic and broad-faced Steppe people).
The Conquest of Western Europe (2500-1200 BCE)
The R1b conquest of Europe happened in two phases. For nearly two millennia, starting from circa 4200 BCE, Steppe people limited their conquest to the rich Chalcolithic civilisations of the Carpathians and the Balkans. These societies possessed the world's largest towns, notably the tell settlements of the Cucuteni-Tripolye culture. Nothing incited the R1b conquerors to move further into Western Europe at such an early stage, because most of the land north and west of the Alps was still sparsely populated woodland. The Neolithic did not reach the British Isles and Scandinavia before circa 4000 BCE. Even northern France and most of the Alpine region had been farming or herding for less than a millennium and were still quite primitive compared to Southeast Europe and the Middle East.
North-west Europe remained a tribal society of hunter-gatherers practising only limited agriculture for centuries after the conquest of the Balkans by the Indo-Europeans. Why would our R1b "conquistadors" leave the comfort of the wealthy and populous Danubian civilisations for the harsh living conditions that lie beyond? Bronze Age people coveted tin, copper, and gold, of which the Balkans had plenty, but that no one had yet discovered in Western Europe.
R1b-L51 is thought to have arrived in Central Europe (Hungary, Austria, Bohemia) around 2500 BCE, approximately two millennia after the shift to the Neolithic lifestyle in these regions. Agrarian towns had started to develop. Gold and copper had begun to be mined. The prospects of a conquest were now far more appealing.
The archeological and genetic evidence (distribution of R1b subclades) point at several consecutive waves towards eastern and central Germany between 2800 BCE and 2300 BCE. The Unetice culture was probably the first culture in which R1b-L11 lineages played a major role. It is interesting to note that the Unetice period happen to correspond to the end of the Maykop (2500 BCE) and Kemi Oba (2200 BCE) cultures on the northern shores of the Black Sea, and their replacement by cultures descended from the northern steppes. It can therefore be envisaged that the (mostly) R1b population from the northern half of the Black Sea migrated westward due to pressure from other Indo-European people (R1a) from the north, for example that of the burgeoning Proto-Indo-Iranian branch, linked to the contemporary Poltavka and Abashevo cultures.
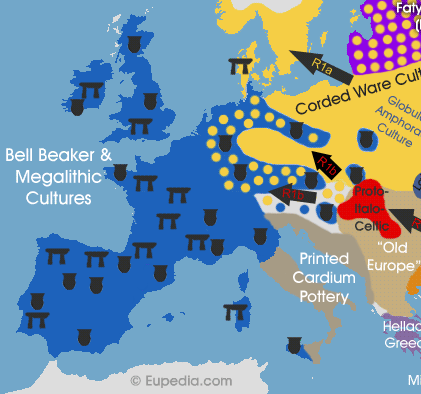
It is doubtful that the Bell Beaker culture (2900-1800 BCE) in Western Europe was already Indo-European because its attributes are in perfect continuity with the native Megalithic cultures. The Beaker phenomenon started during the Late Neolithic and Early Chalcolithic in Portugal and propagated to the north-east towards Germany. During the same period Bronze Age Steppe cultures spread from Germany in the opposite direction towards Iberia, France and Britain, progressively bringing R1b lineages into the Bell Beaker territory. It is more likely that the beakers and horses found across Western Europe during that period were the result of trade with neighbouring Indo-European cultures, including the first wave of R1b into Central Europe. It is equally possible that the Beaker people were R1b merchants or explorers who travelled across Western Europe and brought back tales of riches poorly defended by Stone Age people waiting to be to be conquered. This would have prompted a full-scale Indo-European (R1b) invasion from about 2500 BCE in Germany, reaching the Atlantic (north of the Pyrenees at least) around 2200 BCE.
Ancient DNA tests conducted by Lee et al. (2012), Haak et al. (2015) and Allentoft et al. (2015) have all confirmed the presence of R1b-L51 (and deeper subclades such as P312 and U152) in Germany from the Bell Beaker period onwards, but none in earlier cultures. German Bell Beaker R1b samples only had about 50% of Yamna autosomal DNA and often possessed Neolithic non-Steppe mtDNA, which confirms that R1b invaders took local wives as they advanced westward. Another study by Olalde et al. (2017) confirmed that Iberian Bell Beakers were genetically distinct from the previously tested German samples. None of the Spanish or Portuguese individuals associated with Bell Beaker pottery possessed any Steppe admixture, and none belonged to the Indo-European haplogroup R1b-L23 or its subclades. Instead they belonged to typical Megalithic lineages like G2a, I2a1, I2a2 and the Neolithic R1b-V88. The paper also confirmed a high frequency of R1b-L51 lineages in central Europe during the Beall Beaker period. In Britain, Megalithic individuals belonged exclusively to Y-haplogroup I2 (mostly I2a2 and I2a1b-L161), but were entirely replaced by R1b-L51 (mosly L21 clade) in the Early Bronze Age. This means that the Bell Beaker culture was not associated with one particular ethnic group. Beaker pottery originated in Megalithic Iberia, but then spread to France and central Europe and was used by invading R1b-L51 Steppe people, who brought it with them to the British Isles, while wiping out most of the indigenous Megalithic population. There was therefore no 'Bell Beaker people', but just various populations trading and using Beaker pots during that period.
DNA samples from the Unetice culture (2300-1600 BCE) in Germany, which emerged less than two centuries after the apperance of the first R1b-L51 individuals in the late Bell Beaker Germany, had a slightly higher percentage of Yamna ancestry (60~65%) and of Yamna-related mtDNA lineages, which indicates a migration of both Steppe men and women. That would explain why archeological artefacts from the Unetice culture are clearly Yamna-related (i.e. Indo-European), as they abruptly introduced new technologies and a radically different lifestyle, while the Bell Beaker culture was in direct continuity with previous Neolithic or Chalcolithic cultures. R1b men may simply have conquered the Bell Beaker people and overthrown the local rulers without obliterating the old culture due to their limited numbers. Taking the analogy of the Germanic migrations in the Late Antiquity, the R1b invasion of the Bell Beaker period was more alike to that of the Goths, Burgunds and Vandals, who all migrated in small numbers, created new kingdoms within the Roman empire, but adopted Latin language and Roman culture. In contrast, the Corded Ware and Unetice culture involved large-scale migrations of Steppe people, who imposed their Indo-European language and culture and conquered people, just like the Anglo-Saxons or the Bavarians did in the 5th century.

| Related topics |
|---|
|
|
The cultures that succeeded to Unetice in Central Europe, chronologically the Tumulus culture (1600-1200 BCE), Urnfield culture (1300-1200 BCE) and Hallstatt culture (1200-750 BCE) cultures remained typically Indo-European. The Hallstatt culture, centered around the Alps, is considered the first classical Celtic culture in Europe. It quickly expanded to France, Britain, Iberia, northern Italy and the Danube valley, probably spreading for the first time Celtic languages, although not bronze technology nor R1b lineages, which had both already spread over much of western Europe during the Bell Beaker period.
=> See also Metal-mining and stockbreeding explain R1b dominance in Atlantic fringe
| Did the Indo-Europeans really invade Western Europe? |
|---|
|
Proponents of the Paleolithic or Neolithic continuity model argue that bronze technology and horses could have been imported by Western Europeans from their Eastern European neighbours, and that no actual Indo-European invasion need be involved. It is harder to see how Italic, Celtic and Germanic languages were adopted by Western and Northern Europeans without at least a small scale invasion. It has been suggested that Indo-European (IE) languages simply disseminated through contact, just like technologies, or because it was the language of a small elite and therefore its adoption conferred a certain perceived prestige. However people don't just change language like that because it sounds nicer or more prestigious. Even nowadays, with textbooks, dictionaries, compulsory language courses at school, private language schools for adults and multilingual TV programs, the majority of the people cannot become fluent in a completely foreign language, belonging to a different language family. The linguistic gap between pre-IE vernaculars and IE languages was about as big as between modern English and Chinese. English, Greek, Russian and Hindi are all related IE languages and therefore easier to learn for IE speakers than non-IE languages like Chinese, Arabic or Hungarian. From a linguistic point of view, only a wide-scale migration of IE speakers could explain the thorough adoption of IE languages in Western Europe - leaving only Basque as a remnant of the Neolithic languages.
One important archeological argument in favour of the replacement of Neolithic cultures by Indo-European culture in the Bronze Age comes from pottery styles. The sudden appearance of bronze technology in Western Europe coincides with ceramics suddenly becoming more simple and less decorated, just like in the Pontic Steppe. Until then, pottery had constantly evolved towards greater complexity and details for over 3,000 years. People do not just decide like that to revert to a more primitive style. Perhaps one isolated tribe might experiment with something simpler at one point, but what are the chances that distant cultures from Iberia, Gaul, Italy and Britain all decide to undertake such an improbable shift around the same time? The best explanation is that this new style was imposed by foreign invaders. In this case it is not mere speculation; there is ample evidence that this simpler pottery is characteristic of the steppes associated with the emergence of Proto-Indo-European speakers.
Besides pottery, archaeology provides ample evidence that the early Bronze Age in Central and Western Europe coincides with a radical shift in food production. Agriculture experiences an abrupt reduction in exchange for an increased emphasis on domesticates. This is also a period when horses become more common and cow milk is being consumed regularly. The oeverall change mimicks the Steppic way of life almost perfectly. Even after the introduction of agriculture around 5200 BCE, the Bug-Dniester culture and later Steppe cultures were characterised by an economy dominated by herding, with only limited farming. This pattern expands into Europe exactly at the same time as bronze working.
Religious beliefs and arts undergo a complete reversal in Bronze Age Europe. Neolithic societies in the Near East and Europe had always worshipped female figurines as a form of fertility cult. The Steppe cultures, on the contrary, did not manufacture female figurines. As bronze technology spreads from the Danube valley to Western Europe, symbols of fertility and fecundity progressively disappear and are replaced by scultures of domesticated animals.
Another clue that Indo-European Steppe people came in great number to Central and Western Europe is to be found in burial practices. Neolithic Europeans either cremated their dead (e.g. Cucuteni-Tripolye culture) or buried them in collective graves (this was the case of Megalithic cultures). In the Steppe, each person was buried individually, and high-ranking graves were placed in a funeral chamber and topped by a circular mound. The body was typically accompanied by weapons (maces, axes, daggers), horse bones, and a dismantled wagon (or later chariot). These characteristic burial mounds are known as kurgans in the Pontic Steppe. Men were given more sumptuous tombs than women, even among children, and differences in hierarchy are obvious between burials. The Indo-Europeans had a strongly hierarchical and patrilinear society, as opposed to the more egalitarian and matrilinear cultures of Old Europe. The proliferation of ststus-conscious male-dominant kurgans (or tumulus) in Central Europe during the Bronze Age is a clear sign that the ruling elite had now become Indo-European. The practice also spread to central Asia and southern Siberia, two regions where R1a and R1b lineages are found nowadays, just like in Central Europe. The ceremony of burial is one of the most emotionally charged and personal aspect of a culture. It is highly doubtful that people would change their ancestral practice "just to do like the neighbours". In fact, different funerary practices have co-existed side by side during the European Neolithic and Chalcolithic. The ascendancy of yet another constituent of the Pontic Steppe culture in the rest of Europe, and in this case one that does not change easily through contact with neighbours, adds up to the likelihood of a strong Indo-European migration. The adoption of some elements of a foreign culture tends to happen when one civilization overawes the adjacent cultures by its superiority. This process is called 'acculturation'. However there is nothing that indicates that the Steppe culture was so culturally superior as to motivate a whole continent, even Atlantic cultures over 2000 km away from the Pontic Steppe, to abadndon so many fundamental symbols of their own ancestral culture, and even their own language. In fact, Old Europe was far more refined in its pottery and jewellery than the rough Steppe people. The Indo-European superiority was cultural but military, thanks to horses, bronze weapons and an ethic code valuing individual heroic feats in war (these ethic values are known from the old IE texts, like the Rig Veda, Avesta, or the Mycenaean and Hittite literature).
After linguistics and archaeology, the third category of evidence comes from genetics itself. It had first been hypothetised that R1b was native to Western Europe, because this is where it was most prevalent. It has since been proven that R1b haplotypes displayed higher microsatellite diversity in Anatolia and in the Caucasus than in Europe. European subclades are also more recent than Middle Eastern or Central Asian ones. The main European subclade, R-P312/S116, only dates back to approximately 3500 to 3000 BCE. It does not mean that the oldest common ancestor of this lineage arrived in Western Europe during this period, but that the first person who carried the mutation R-P312/S116 lived at least 5,000 years ago, assumably somewhere in the lower Danube valley or around the Black Sea. In any case this timeframe is far too recent for a Paleolithic origin or a Neolithic arrival of R1b. The discovery of what was thought to be "European lineages" in Central Asia, Pakistan and India hit the final nail on the coffin of a Paleolithic origin of R1b in Western Europe, and confirmed the Indo-European link.
All the elements concur in favour of a large scale migration of Indo-European speakers (possibly riding on horses) to Western Europe between 2500 to 2100 BCE, contributing to the replacement of the Neolithic or Chalcolithic lifestyle by a inherently new Bronze Age culture, with simpler pottery, less farming, more herding, new rituals (single graves) and new values (patrilinear society, warrior heroes) that did not evolve from local predecessors.
|
The Atlantic Celtic branch (L21)
The Proto-Italo-Celto-Germanic R1b people had reached in what is now Germany by 2500 BCE. By 2300 BCE they had arrived in large numbers and founded the Unetice culture. Judging from the propagation of bronze working to Western Europe, those first Indo-Europeans reached France and the Low Countries by 2200 BCE, Britain by 2100 BCE and Ireland by 2000 BCE, and Iberia by 1800 BCE. This first wave of R1b presumably carried R1b-L21 lineages in great number (perhaps because of a founder effect), as these are found everywhere in western, northern and Central Europe. Cassidy et al. (2015) confirmed the presence of R1b-L21 (DF13 and DF21 subclades) in Ireland around 2000 BCE. Those genomes closely resembled those of the Unetice culture autosomally, but differed greatly from the earlier Neolithic Irish samples. This confirms that a direct migration of R1b-L21 from Central Europe was responsible for the introduction of the Bronze Age to Ireland.
The early split of L21 from the main Proto-Celtic branch around Germany would explain why the Q-Celtic languages (Goidelic and Hispano-Celtic) diverged so much from the P-Celtic branch (La Tène, Gaulish, Brythonic), which appears to have expanded from the later Urnfield and Hallstat cultures.
Some L21 lineages from the Netherlands and northern Germany later entered Scandinavia (from 1700 BCE) with the dominant subclade of the region, R1b-S21/U106 (see below). The stronger presence of L21 in Norway and Iceland can be attributed to the Norwegian Vikings, who had colonised parts of Scotland and Ireland and taken slaves among the native Celtic populations, whom they brought to their new colony of Iceland and back to Norway. Nowadays about 20% of all Icelandic male lineages are R1b-L21 of Scottish or Irish origin.
In France, R1b-L21 is mainly present in historical Brittany (including Mayenne and Vendée) and in Lower Normandy. This region was repopulated by massive immigration of insular Britons in the 5th century due to pressure from the invading Anglo-Saxons. However, it is possible that L21 was present in Armorica since the Bronze age or the Iron age given that the tribes of the Armorican Confederation of ancient Gaul already had a distinct identity from the other Gauls and had maintained close ties with the British Isles at least since the Atlantic Bronze Age.
Distribution of haplogroup R1b-L21 (S145) in Europe
 in Europe")
The Gallic & Iberian branch (DF27/S250)
The first Proto-Celtic R1b lineages to reach France and the Iberian peninsula from Central Europe were probably L21 and DF27. Whereas L21 might have taken a northern route through Belgium and northern France on its way to the British Isles, DF27 seems to have spread all over France but heading in greater number toward the south.
The Bronze Age did not appear in Iberia until 1800 BCE, and was mostly confined to the cultures of El Argar and Los Millares in south-east Spain, with sporadic sites showing up in Castile by 1700 BCE and in Extremadura and southern Portugal by 1500 BCE. These Early Bronze Age sites typically did not have more than some bronze daggers or axes and cannot be considered proper Bronze Age societies, but rather Copper Age societies with occasional bronze artefacts (perhaps imported). These cultures might have been founded by small groups of R1b adventurers looking for easy conquests in parts of Europe that did not yet have bronze weapons. They would have become a small ruling elite, would have had children with local women, and within a few generations their Indo-European language would have been lost, absorbed by the indigenous languages (=> see How did the Basques become R1b?).
Martiniano et al. (2017) sequenced the genomes of various skeletons from West Iberia dating from the Middle and Late Neolithic, Chalcolithic and Middle Bronze Age (since the Early Bronze Age did not reach that region). They found that Neolithic and Chalcolithic individuals belonged to Y-haplogroups I*, I2a1 and G2a. In contrast, all three Bronze Age Portuguese men tested belonged to R1b (one M269 and two P312), although they carried Neolithic Iberian maternal lineages (H1, U5b3, X2b) and lacked any discernible Steppe admixture. This is concordant with a scenario of Indo-European R1b men entering Iberia from 1800 BCE as a small group of adventurers and taking local wives, thus diluting their DNA at each generation, until hardly any Steppe admixture was left after a few centuries, by the time they reached Portugal. Nowadays, Spaniards and Portuguese do possess about 25% of Steppe admixture, which means that other more important Indo-European migrations took place later on, during the Late Bronze Age and the Iron Age.
Iberia did not become a fully-fledged Bronze Age society until the 13th century BCE, when the Urnfield culture (1300-1200 BCE) expanded from Germany to Catalonia via southern France, then the ensuing Hallstatt culture (1200-750 BCE) spread throughout most of the peninsula (especially the western half). This period belongs to the wider Atlantic Bronze Age (1300-700 BCE), when Iberia was connected to the rest of Western Europe through a complex trade network.
It is hard to say when exactly DF27 entered Iberia. Considering its overwhelming presence in the peninsula and in south-west France, it is likely that DF27 arrived early, during the 1800 to 1300 BCE period, and perhaps even earlier, if R1b adventurers penetrated the Bell Beaker culture, as they appear to have done all over Western Europe from 2300 BCE to 1800 BCE. The Atlantic Bronze Age could correspond to the period when DF27 radiated more evenly around Iberia and ended up, following Atlantic trade routes, all the way to the British Isles, the Netherlands and Scandinavia.
Distribution of haplogroup R1b-DF27 in Europe

The Italo-Celtic branch (S28/U152/PF6570)
Furtwängler et al. (2020) analysed 96 ancient genomes from Switzerland, Southern Germany, and the Alsace region in France, covering the Middle/Late Neolithic to Early Bronze Age. They confirmed that R1b arrived in the region during the transitory Bell Beaker period (2800-1800 BCE). The vast majority of Bell Beaker R1b samples belonged to the U152 > L2 clade (11 out of 14; the other being P312 or L51).
Starting circa 1300 BCE, a new Bronze Age culture flourished around the Alps thanks to the abundance of metal in the region, and laid the foundation for the classical Celtic culture. It was actually the succession of three closely linked culture: the Urnfield culture, which would evolve into the Hallstatt culture (from 1200 BCE) and eventually into the La Tène culture (from 450 BCE). After the Unetice expansion to Western Europe between 2300 and 1800 BCE, the Urnfield/Hallstatt/La Tène period represents the second major R1b expansion that took place from Central Europe, pushing west to the Atlantic, north to Scandinavia, east to the Danubian valley, and eventually as far away as Greece, Anatolia, Ukraine and Russia, perhaps even until the Tarim basin in north-west China (=> see Tarim mummies.
R1b-U152 would have entered Italy in successive waves from the northern side of the Alps, starting in 1700 BCE with the establishment of the Terramare culture in the Po Valley. From 1200 BCE, a larger group of Hallstatt-derived tribes founded the Villanova culture (see below). This is probably the migration that brought the Italic-speaking tribes to Italy, who would have belonged mainly the Z56 clade of R1b-U152. During the Iron Age, the expansion of the La Tène culture from Switzerland is associated with the diffusion of the Z36 branch, which would generate the Belgae around modern Belgium and in the Rhineland, the Gauls in France, and the Cisalpine Celts in Italy.
Antonio et al. (2019) analysed the genomes of Iron Age Latins dating between 900 and 200 BCE, and the samples tested belonged primarily to haplogroup R1b-U152 (including the clades L2, Z56 and Z193), as well as one R1b-Z2103 and one R1b-Z2118.

One common linguistic trait between Italic and Gaulish/Brythonic Celtic languages linked to the Hallstatt expansion is that they shifted the original IE *kw sound into *p. They are known to linguists as the P-Celtic branch (as opposed to Q-Celtic). It is thought that this change occured due to the inability to pronounce the *kw sound by the pre-Indo-European population of Central Europe, Gaul and Italy, who were speakers of Afro-Asiatic dialects that had evolved from Near-Eastern languages inherited from the Neolithic. The Etruscans, although later incomers from the eastern Mediterranean, also fit in this category. It has recently been acknowledged that Celtic languages borrowed part of their grammar from Afro-Asiatic languages. This shift could have happened when the Proto-Italo-Celtic speakers moved from the steppes to the Danube basin and mixed with the population of Near-Eastern farmers belonging to haplogroups E1b1b, G2a, J and T. However, such an early shift would not explain why Q-Celtic and Germanic languages did not undergo the same linguistic mutation. It is therefore more plausible that the shift happened after the Proto-Italo-Celts and Proto-Germanics had first expanded across all western and northern Europe. The S28/U152 connection to P-Celtic (and Italic) suggests that the shift took place around the Alps after 1800 BCE, but before the invasion of Italy by the Italic tribes circa 1200 BCE.
The expansion of the Urnfield/Hallstatt culture to Italy is evident in the form of the Villanovan culture (c. 1100-700 BCE), which shared striking resemblances with the Urnfield/Hallstatt sites of Bavaria and Upper Austria. The Villanova culture marks a clean break with the previous Terramare culture. Although both cultures practised cremation, whereas Terramare people placed cremated remains in communal ossuaries like their Neolithic ancestors from the Near East, Villanovans used distinctive Urnfield-style double-cone shaped funerary urns, and elite graves containing jewellery, bronze armour and horse harness fittings were separated from ordinary graves, showing for the first time the development of a highly hierarchical society, so characteristic of Indo-European cultures. Quintessential Indo-European decorations, such as swastikas, also make their appearance. Originally a Bronze-age culture, the Villanova culture introduced iron working to the Italian peninsula around the same time as it appeared in the Hallstatt culture, further reinforcing the link between the two cultures. In all likelihood, the propagation of the Villanova culture represents the Italic colonisation of the Italian peninsula. The highest proportion of R1b-U152 is found precisely where the Villanovans were the more strongly established, around modern Tuscany and Emilia-Romagna. The Villanova culture was succeeded by the Etruscan civilisation, which displayed both signs of continuity with Villanova and new hybrid elements of West Asian origins, probably brought by Anatolian settlers (who would have belonged to a blend of haplogroups G2a, J2 and R1b-Z2103).
Distribution of haplogroup R1b-S28 (U152) in Europe
 in Europe")
The Germanic branch (S21/U106/M405)
The principal Proto-Germanic branch of the Indo-European family tree is R1b-S21 (a.k.a. U106 or M405). This haplogroup is found at high concentrations in the Netherlands and north-west Germany. It is likely that R1b-S21 lineages expanded in this region through a founder effect during the Unetice period, then penetrated into Scandinavia around 1700 BCE (probably alongside R1a-L664), thus creating a new culture, that of the Nordic Bronze Age (1700-500 BCE). R1b-S21 would then have blended for more than a millennium with preexisting Scandinavian populations, represented by haplogroups I1, I2-L801, R1a-Z284. When the Germanic Iron Age started c. 500 BCE, the Scandinavian population had developed a truly Germanic culture and language, but was divided in many tribes with varying levels of each haplogroup. R1b-S21 became the dominant haplogroup among the West Germanic tribes, but remained in the minority against I1 and R1a in East Germanic and Nordic tribes, including those originating from Sweden such as the Goths, the Vandals and Lombards.
The presence of R1b-S21 in other parts of Europe can be attributed almost exclusively to the Germanic migrations that took place between the 3rd and the 10th century. The Frisians and Anglo-Saxons disseminated this haplogroup to England and the Scottish Lowlands, the Franks to Belgium and France, the Burgundians to eastern France, the Suebi to Galicia and northern Portugal, and the Lombards to Austria and Italy. The Goths help propagate S21 around Eastern Europe, but apparently their Germanic lineages were progressively diluted by blending with Slavic and Balkanic populations, and their impact in Italy, France and Spain was very minor. Later the Danish and Norwegian Vikings have also contributed to the diffusion of R1b-S21 (alongside I1, I2b1 and R1a) around much of Western Europe, but mainly in Iceland, in the British Isles, in Normandy, and in the southern Italy.
From the Late Middle Ages until the early 20th century, the Germans expanded across much of modern Poland, pushing as far as Latvia to the north-east and Romania to the south-east. During the same period the Austrians built an empire comprising what is now the Czech Republic, Slovakia, Hungary, Slovenia, Croatia, Serbia, and parts of Romania, western Ukraine and southern Poland. Many centuries of German and Austrian influence in central and Eastern Europe resulted in a small percentage of Germanic lineages being found among modern populations. In Romania 4% of the population still consider themselves German. The low percentage of R1b-S21 in Finland, Estonia and Latvia can be attributed to the Swedish or Danish rule from the late Middle Ages to the late 19th century.
O'Sullivan et al. (2018) tested the genomes of Merovingian nobles from an early Medieval Alemannic graveyard in Baden-Württemberg. Apart from one individual belonging to haplogroup G2a2b1, all men were members of R1b, and all samples that yielded deep clade results fell under the R1b-U106 > Z381 > Z301 > L48 > Z9 > Z325 clade. The lineage of the Kings of France was inferred from the Y-DNA of several descendant branches (see famous members below) and also belongs to R1b-U106 > Z381. Their earliest-known male-line ancestor was from Robert II, Count of Hesbaye, a Frankish nobleman from present-day Belgium. The House of Wettin (see famous members below), one of the oldest dynasties in Europe, which ruled over many states at various times in history, was yet another well-known noble Germanic lineage part of R1b-U106 > Z381.
Distribution of haplogroup R1b-S21 (U106) in Europe
 in Europe")
| How did R1b come to replace most of the older lineages in Western Europe? |
|---|
|
Until recently it was believed that R1b originated in Western Europe due to its strong presence in the region today. The theory was that R1b represented the Paleolithic Europeans (Cro-Magnon) that had sought refuge in the Franco-Cantabrian region at the peak of the last Ice Age, then recolonised Central and Northern Europe once the ice sheet receded. The phylogeny of R1b proved that this scenario was not possible, because older R1b clades were consistently found in Central Asia and the Middle East, and the youngest in Western and Northern Europe. There was a clear gradient from East to West tracing the migration of R1b people (see map above). This age of the main migration from the shores of the Black Sea to Central Europe also happened to match the timeframe of the Indo-European invasion of Europe, which coincides with the introduction of the Bronze-Age culture in Western Europe, and the proliferation of Italo-Celtic and Germanic languages.
Historians and archeologists have long argued whether the Indo-European migration was a massive invasion, or rather a cultural diffusion of language and technology spread only by a small number of incomers. The answer could well be "neither". Proponents of the diffusion theory would have us think that R1b is native to Western Europe, and R1a alone represent the Indo-Europeans. The problem is that haplogroup R did arise in Central Asia, and R2 is still restricted to Central and South Asia, while R1a and the older subclades of R1b are also found in Central Asia. The age of R1b subclades in Europe coincide with the Bronze-Age. R1b must consequently have replaced most of the native Y-DNA lineages in Europe from the Bronze-Age onwards.
However, a massive migration and nearly complete anihilation of the Paleolithic population can hardly be envisaged. Western Europeans do look quite different in Ireland, Holland, Aquitaine or Portugal, despite being all regions where R1b is dominant. Autosomal DNA studies have confirmed that the Western European population is far from homogeneous. A lot of maternal lineages (mtDNA) also appear to be of Paleolithic origin (e.g. H1, H3, U5 or V) based on ancient DNA tests. What a lot of people forget is that there is also no need of a large-scale exodus for patrilineal lineages to be replaced fairly quickly. Here is why.
- Polygamy. Unlike women, men are not limited in the number of children they can procreate. Men with power typically have more children. This was all the truer in primitive societies, where polygamy was often the norm for chieftains and kings.
- Status & Power. Equipped with Bronze weapons and horses, the Indo-Europeans would have easily subjugated the Neolithic farmers and with even greater ease Europe's last hunter-gatherers.If they did not exterminate the indigenous men, the newcomers would have become the new ruling class, with a multitude of local kings, chieftains and noblemen (Bronze-Age Celts and Germans lived in small village communities with a chief, each part of a small tribe headed by a king) with higher reproductive opportunities than average.
- Gender imbalance. Invading armies normally have far more men than women. Men must therefore find women in the conquered population. Wars are waged by men, and the losers suffer heavier casualties, leaving more women available to the winners.
- Aggressive warfare. The Indo-Europeans were a warlike people with a strong heroic code emphasising courage and military prowess. Their superior technology (metal weapons, wheeled vehicles drawn by horses) and attitude to life would have allowed them to slaughter any population that did not have organised armies with metal weapons (i.e. anybody except the Middle-Eastern civilizations).
- Genetic predisposition to conceive boys. The main role of the Y-chromosome in man's body is to create sperm. Haplogroups are determined based on mutations differentiating Y-chromosomes. Each mutation is liable to affect sperm production and sperm motility. Preliminary research has already established a link between certain haplogroups and increased or reduced sperm motility. The higher the motility, the higher the chances of conceiving a boy. It is absolutely possible that R1b could confer a bias toward more male offspring. Even a slightly higher percentage of male births would significantly contribute to the replacement of other lineages with the accumulation effect building up over a few millennia. Not all R1b subclades might have this boy bias. The bias only exist in relation to other haplogroups found in a same population. It is very possible that the fairly recent R1b subclades of Western Europe had a significant advantage compared to the older haplogroups in that region, notably haplogroup I2 and E-V13. Read more
Replacement of patrilineal lineages following this model quickly becomes exponential. Imagine 100 Indo-European men conquering a tribe of 1000 indigenous Europeans (a ratio of 1:10). War casualties have resulted in a higher proportion of women in the conquered population. Let's say that the surviving population is composed of 700 women and 300 men. Let's suppose that the victorious Indo-European men end up having twice as many children reaching adulthood as the men of the vanquished tribe. There is a number of reason for that. The winners would take more wives, or take concubines, or even rape women of the vanquished tribe. Their higher status would garantee them greater wealth and therefore better nutrition for their offspring, increasing the chances of reaching adulthood and procreating themselves. An offspring ratio of 2 to 1 for men is actually a conservative estimate, as it is totally conceivable that Bronze-Age sensibilities would have resulted in killing most of the men on the losing side, and raping their women (as attested by the Old Testament). Even so, it would only take a few generations for the winning Y-DNA lineages to become the majority. For instance, if the first generation of Indo-Europeans had two surviving sons per man, against only one per indigenous man, the number of Indo-European paternal lineages would pass to 200 individuals at the second generation, 400 at the third, 800 at the fourth and 1600 at the fifth, and so on. During that time indigenous lineages would only stagnate at 300 individuals for each generation.
Based on such a scenario, the R1b lineages would have quickly overwhelmed the local lineages. Even if the Indo-European conquerors had only slightly more children than the local men, R1b lineages would become dominant within a few centuries. Celtic culture lasted for over 1000 years in Continental Europe before the Roman conquest putting an end to the priviledges of the chieftains and nobility. This is more than enough time for R1b lineages to reach 50 to 80% of the population.
The present-day R1b frequency forms a gradient from the Atlantic fringe of Europe (highest percentage) to Central and Eastern Europe (lowest), the rises again in the Anatolian homeland. This is almost certainly because agriculture was better established in Eastern, then Central Europe, with higher densities of population, leaving R1b invadors more outnumbered than in the West. Besides, other Indo-Europeans of the Corded Ware culture (R1a) had already advanced from modern Russia and Ukraine as far west as Germany and Scandinavia. It would be difficult for R1b people to rival with their R1a cousins who shared similar technology and culture. The Pre-Celto-Germanic R1b would therefore have been forced to settled further west, first around the Alps, then overtaking the then sparsely populated Western Europe.
|
The Balkanic and Asian branch (Z2103)
Asian branch
Haak et al. (2015) tested six Y-DNA samples from the eastern reaches of Yamna culture, in the Volga-Ural region, and all of them turned out to belong to haplogroup R1b. Four of them were positive for the Z2103 mutation. IN all likelihood, R1b-Z2103 was a major lineage of the Poltavka culture, which succeeded to the Yamna culture between the Volga River and the Ural mountains. It eventually merged with the Abashevo culture (presumably belonging chiefly to R1a-Z93) to form the Sintashta culture. Through a founder effect or through political domination, R1a-Z93 lineages would have outnumbered R1b-Z2103 after the expansion to Central and South Asia, although important pockets of Z2103 survived, notably in Bashkorostan, Turkmenistan and Uyghurstan (Chinese Turkestan).
R1b-Z2103 would have become an Indo-Iranian lineage like R1a-Z93. This is true of two Z2103 subclades in particular: L277.1 and L584. The former is found in Russia to Central Asia then to India and the Middle East, just like the R1a-L657 subclade of Z93. It can be associated with the Andronovo culture and BactriaMargiana Archaeological Complex, as well as the Indo-Aryan migrations. R1b-L584 is found especially in Iran, northern Iraq, the South Caucasus and Turkey, and correlates more with the Iranian branch of Indo-Europeans, which includes Persians, Kurds and Scythians.
Distribution of haplogroup R1b-ht35 (Z2103) in Europe
 in Europe")
Anatolian branch
The Hittites (c. 2000-1178 BCE) were the first Indo-Europeans to defy (and defeat) the mighty Mesopotamian and Egyptian empires. There are two hypotheses regarding the origins of the Hittites. The first is that they came from the eastern Balkans and invaded Anatolia by crossing the Bosphorus. That would mean that they belonged either to the L23* or the Z2103 subclade. The other plausible scenario is that they were an offshoot of the late Maykop culture, and that they crossed the Caucasus to conquer the Hattian kingdom (perhaps after being displaced from the North Caucasus by the R1a people of the Catacomb culture). In that case the Hittites might have belonged to the R1b-Z2103 or the R1b-PF7562 subclade. The first hypothesis has the advantage of having a single nucleus, the Balkans, as the post-Yamna expansion of all Indo-European R1b. The Maykop hypothesis, on the other hand, would explain why the Anatolian branch of IE languages (Hittite, Luwian, Lydian, Palaic) is so archaic compared to other Indo-European languages, which would have originated in Yamna rather than Maykop.
There is substantial archaeological and linguistic evidence that Troy was an Indo-European city associated with the Steppe culture and haplogroup R1b. The Trojans were Luwian speakers related to the Hittites (hence Indo-European), with attested cultural ties to the culture of the Pontic-Caspian Steppe. The first city of Troy dates back to 3000 BCE, right in the middle of the Maykop period. Troy might have been founded by Maykop people as a colony securing the trade routes between the Black Sea and the Aegean. The founding of Troy happens to coincide exactly with the time the first galleys were made. Considering the early foundation of Troy, the most likely of the two Indo-European paternal haplogroups would be R1b-M269 or L23.
The Phrygians and the Proto-Armenians are two other Indo-European tribes stemming from the Balkans. Both appear to have migrated to Anatolia around 1200 BCE, during the 'great upheavals' of the Eastern Mediterranean (see below). The Phrygians (or Bryges) founded a kingdom (1200-700 BCE) in west central Anatolia, taking over most of the crumbling Hittite Empire. The Armenians crossed all Anatolia until Lake Van and settled in the Armenian Highlands. Nowadays 30% of Armenian belong to haplogroup R1b, the vast majority to the L584 subclade of Z2103 (=> see The Indo-European migrations to Armenia).
Most of the R1b found in Greece today is of the Balkanic Z2103 variety. There is also a minority of Proto-Celtic S116/P312 and of Italic/Alpine Celtic S28/U152. Z2103 could have descended from Albania or Macedonia during the Dorian invasion (see below), thought to have happened in the 12th century BCE. Their language appear to have been close enough to Mycenaean Greek to be mutually intelligible and easy for locals to adopt. The Mycenaeans might have brought some R1b (probably also Z2103) to Greece, but their origins can be traced back through archaeology to the Catacomb culture and the Seima-Turbino phenomenon of the northern forest-steppe, which would make them primarily an R1a tribe.
Greek and Anatolian S116 and some S28 lineages could be attributed to the La Tène Celtic invasions of the 3rd century BCE. The Romans also certainly brought S28 lineages (=> see Genetics of the Italian people), and probably also the Venetians later on, notably on the islands. Older clades of R1b, such as P25 and V88, are only a small minority and would have come along E1b1b, G2a and J2 from the Middle East.
| The great upheavals circa 1200 BCE |
|---|
|
1200 BCE was a turning point in European and Near-Eastern history. In Central Europe, the Urnfield culture evolved into the Hallstatt culture, traditionally associated with the classical Celtic civilization, which was to have a crucial influence on the development of ancient Rome. In the Pontic Steppe, the Srubna culture make way to the Cimmerians, a nomadic people speaking an Iranian or Thracian language. The Iron-age Colchian culture (1200-600 BCE) starts in the North Caucasus region. Its further expansion to the south of the Caucasus correspond to the first historical mentions of the Proto-Armenian branch of Indo-European languages (circa 1200 BCE). In the central Levant the Phoenicians start establishing themselves as significant maritime powers and building their commercial empire around the southern Mediterranean.
But the most important event of the period was incontestably the destruction of the Near-Eastern civilizations, possibly by the Sea Peoples. The great catastrophe that ravaged the whole Eastern Mediterranean from Greece to Egypt circa 1200 BCE is a subject that remains controversial. The identity of the Sea Peoples has been the object of numerous speculations. What is certain is that all the palace-based societies in the Near-East were abruptly brought to an end by tremendous acts of destruction, pillage and razing of cities. The most common explanation is that the region was invaded by technologically advanced warriors from the north. They could have been either Indo-Europeans descended from the Steppe via the Balkans, or Caucasian people (G2a, J1, J2a, T1a) linked with the expansion of the earlier Kura-Araxes culture to eastern Anatolia and the Levant.
The Hittite capital Hattusa was destroyed in 1200 BCE, and by 1160 BCE the empire had collapsed, probably under the pressure of the Phrygians and the Armenians coming from the Balkans. The Mycenaean cities were ravaged and abandoned throughout the 12th century BCE, leading to the eventual collapse of Mycenaean civilization by 1100 BCE. The kingdom of Ugarit in Syria was annihilated and its capital never resettled. Other cities in the Levant, Cyprus and Crete were burned and left abandoned for many generations. The Egyptians had to repel assaults from the Philistines from the East and the Libyans from the West - two tribes of supposed Indo-European origin. The Lybians were accompanied by mercenaries from northern lands (the Ekwesh, Teresh, Lukka, Sherden and Shekelesh), whose origin is uncertain, but has been placed in Anatolia, Greece and/or southern Italy.
The devastation of Greece followed the legendary Trojan War (1194-1187 BCE). It has been postulated that the Dorians, an Indo-European people from the Balkans (probably coming from modern Bulgaria or Macedonia), invaded a weakened Mycenaean Greece after the Trojan War, and finally settled in Greece as one of the three major ethnic groups. The Dorian regions of classical Greece, where Doric dialects were spoken, were essentially the southern and eastern Peloponnese, Crete and Rhodes, which is also the part of Greece with the highest percentage of R1b-Z2103.
Another hypothesis is that the migration of the Illyrians from north-east Europe to the Balkans displaced previous Indo-European tribes, namely the Dorians to Greece, the Phrygians to north-western Anatolia and the Libu to Libya (after a failed attempt to conquer the Nile Delta in Egypt). The Philistines, perhaps displaced from Anatolia, finally settled in Palestine around 1200 BCE, unable to enter Egypt.
|
Other migrations of R1b
Other migrations occured from Europe to the Near East and Central Asia during the Antiquity and Middle Ages. R1b-S28 (U152) was found in Romania, Turkey, northern Bashkortostan (a staggering 71.5% of the local population according to Myres et al.), and at the border of Kazakhstan and Kyrgyzstan. Some of it was surely brought by the La Tène Celts, known to have advanced along the Danube, and created the Galatian kingdom in central Anatolia. The rest could just as well be Roman, given that R1b-S28 is the dominant form of R1b in the Italian peninsula. Some have hypothetised that some "lost" Roman legions went as far as Central Asia or China and never came back, marrying local women and leaving their genetic marker in isolated pockets in Asia. A more prosaic version is that Roman merchants ended up in China via the Silk Road, which existed since the 2nd century BCE.
A small percentage of Western European R1b subclades were also found among Christian communities in Lebanon. They are most likely descendants of the crusaders.
The lactase persistence allele and R1b cattle pastoralists
Lactose (milk sugar) is an essential component of breast milk consumed by infants. Its digrestion is made possible by an enzyme, called lactase, which breaks down lactose in simple sugars that can be absorbed through the intestinal walls and into the bloodstream. In most mammals (humans included), the production of the lactase enzyme is dramatically reduced soon after weaning. As a result, older children and adults become lactose intolerant. That is true of a big part of the world population. Some people possess a genetic mutation that allows the production of lactase through adulthood. This is called lactase persistence (LP). Lactase persistence is particularly common among Northwest Europeans, descended from the ancient Celtic and Germanic people, and in parts of Africa where cattle herding has been practiced for thousands of years. The highest incidence for the lactase persistence alleles, known to geneticists as -13,910*T (rs4988235) and -22018*A (rs182549), are found among Scandinavian, Dutch, British, Irish and Basque people. Sub-Saharan populations with lactase persistence have different mutations, such as -14010*C, -13915*G and -13907*G.
R1b men are thought to be the first people on earth to successfully domesticate cattle and to develop a lifestyle based on cattle husbandry and herding during the Pre-Pottery Neolithic (see Neolithic section). Looking for pasture for their cows, R1b tribes migrated from the Near East to the savannah of North Africa (which has since underwent desertification and become the Sahara) and to the Pontic Steppe in southern Russia and Ukraine. For several millennia no other human population was so depended on cattle for their survival as these R1b tribes.
It is known that most Neolithic herding societies consumed at least some animal milk and even made cheese from it (since cheese contains less lactose and is easier to digest for people who are lactose intolerant). In most of Europe, the Middle East and South Asia, people essentially herded goats and sheep, better suited to mountainous environment of the Mediterranean basin, Anatolia and Iran. Goats and sheep could also be kept easily inside villages by sedentary cereal cultivators, while cows needed vast pastures for grazing, which were particularly scare in the Middle East. Domesticated cattle were sometimes found in small number among other Neolithic populations, but the ones that relied almost entirely on them were the R1b tribes of the Pontic Steppe and North Africa. To this very day, semi-nomadic pastoralists in the Sahel, such as the Fulani and the Hausa, who are descended from Neolithic R1b-V88 migrants from the Near East, still maintain primarily herds of cattle. It is among these cattle herders that selective pressure for lactase persistence would have been the strongest.
There has been speculations among geneticists and evolutionary biologists regarding the origin of the lactase persistence allele in Europeans. Over 100 ancient DNA samples have been tested from Mesolithic, Neolithic and Bronze Age Europe and Syria, and the -13910*T allele has been found only in Late Neolithic/Chalcolithic and Bronze Age individuals. The origin of the mutation does not really matter, since it could have been present at low frequencies in the human gene pool for tens of thousands of years before it underwent postive selective pressure among cattle-herding societies. What is certain is that individuals from Bronze Age cultures associated with the arrival of Indo-European speakers from the Pontic Steppe already possessed relatively high percentages of the LP allele. For example the LP allele was found at a frequency of 27% (see Schilz 2006) among the 13 individuals from the Lichtenstein Cave in Germany, who belonged to the Urnfield culture, and were a mix of Y-haplogroups R1b, R1a and I2a2b.
Nowadays, the LP allele is roughly proportional to the percentage of R1b, and to a lower extent R1a, found in a population. In the British Isles, the Low Countries and south-west Scandinavia, where LP is the highest in the world, the combined percentage of R1a and R1b exceeds 70% of the population. In Iberia, the highest percentage of LP is observed among the Basques, who have the highest percentage of R1b. In Italy, LP is most common in the north, like R1b. The lowest incidence of LP in Europe are found in South Italy, Greece and the Balkans, the regions that have the least R1b lineages.
Tishkoff et al. (2017) confirmed that the Hausa and the Fulani, two Sahel tribes with high incidence of R1b-V88, possessed the same LP allele as Europeans, but that East African pastoralist populations with a high prevalence of the lactase persistence trait possess a completely different mutation, which arose independently. This finding is the strongest evidence so far that the -13,910*T allele originated with the first R1b cattle herders in the Near East, who are the ancestors of both the Indo-Europeans and of African R1b-V88 tribes.
R1 populations spread genes for light skin, blond hair and red hair
There is now strong evidence that both R1a and R1b people contributed to the diffusion of the A111T mutation of the SLC24A5, which explains apporximately 35% of skin tone difference between Europeans and Africans, and most variations within South Asia. The distribution pattern of the A111T allele (rs1426654) of matches almost perfectly the spread of Indo-European R1a and R1b lineages around Europe, the Middle East, Central Asia and South Asia. The mutation was probably passed on in the Early neolithic to other Near Eastern populations, which explains why Neolithic farmers in Europe already carried the A111T allele (e.g. Keller 2012 p.4, Lazaridis 2014 suppl. 7), although at lower frequency than modern Europeans and southern Central Asians.
The light skin allele is also found at a range of 15 to 30% in in various ethnic groups in northern sub-Saharan Africa, mostly in the Sahel and savannah zones inhabited by tribes of R1b-V88 cattle herders like the Fulani and the Hausa. This would presuppose that the A111T allele was already present among all R1b people before the Pre-Pottery Neolithic split between V88 and P297. R1a populations have an equally high incidence of this allele as R1b populations. On the other hand, the A111T mutation was absent from the 24,000-year-old R* sample from Siberia, and is absent from most modern R2 populations in Southeast India and Southeast Asia. Consequently, it can be safely assumed that the mutation arose among the R1* lineage during the late Upper Paleolithic, probably some time between 20,000 and 13,000 years ago.
Fair hair was another physical trait associated with the Indo-Europeans. In contrast, the genes for blue eyes were already present among Mesolithic Europeans belonging to Y-haplogroup I. The genes for blond hair are more strongly correlated with the distribution of haplogroup R1a, but those for red hair have not been found in Europe before the Bronze Age, and appear to have been spread primarily by R1b people (=> see The origins of red hair).
The maternal lineages (mtDNA) corresponding to haplogroup R1b
Identifying mtDNA lineages spread by the Indo-European migrations
=> See the main article:
What were the original mtDNA lineages of Neolithic R1b tribes in the Near East?
R1b tribes are thought to have domesticated cattle in that region 10,500 years ago, yet only moved across the Caucasus some time between 7,500 and 6,500 years ago. For three or four millennia, semi-nomadic R1b herders were bound to have intermingled with some of the Near Eastern or Caucasian neighbours. One way of determining what mt-haplogroups R1b tribes carried at the very beginning of the Neolithic, is to compare the above haplogroups with those of African ethnic groups known to possess elevated percentages of R1b-V88. The best studied group are the Fulani, whose mtDNA include three European-looking haplogroups H, J1b1a, U5 and V making up about 15% of their total maternal lineages. These haplogroups have been identified in all four Central African countries sampled, confirming a strong correlation with haplogroup R1b. However, their H, V and U5 could have come from the Berbers of Northwest Africa. The Berbers also carry R1b-V88, but it's possible that some of it came from different Neolithic migrations, including a re-expansion from Iberia, as Berbers carry H1, H3, V1a1a, V5 and U5b1b1, lineages that are all found in the Iberian peninsula. U5b1b1 descends from Mesolithic West Europeans, but at present it is not yet clear how the other haplogroups reached Iberia or Northwest Africa. One hypothesis is that they came from the Near East during the Neolithic, perhaps with R1b-V88 tribes.
African R1b-V88 and Eurasian haplogroup R1b-P297 split roughly 10,000 years ago, almost certainly in Eastern Europe, where they carried mostly mt-haplogroup U5. Toward the end of the last glaciation, some R1b men would have migrated from Eastern Europe to the region of modern Kurdistan accompanied by women belonging to mtDNA U5. Soon after they arrived J1b1a (and maybe V) would have been the first indigenous Near Eastern lineages assimilated by R1b tribes. R1b-V88 might have assimilated H1 and H3 women in the Levant before moving to North Africa, but that remains highly hypothetical.
Famous individuals
R1b-S1200 branch

The Adams Surname Y-DNA Project compared the Y-DNA of 15 families that have an oral history of being related to the US Presidents John Adams (second president of the United States) and his son John Quincy Adams (6th president). All the men tested share the same haplotype that falls into haplogroup R1b-S1200 > S14328 > S1196 > S6868 > FGC23892. The statesman, political philosopher, and Founding Father Samuel Adams (1722-1803) also shares the same paternal lineage.

The great American inventor Thomas Alva Edison (1847-1931) appears to have belonged to R1b-S1200 > S14328 > SZ5489 > Y28597 > S10242 according to the Edison Y-DNA project (lineage 1). Edison was of one of the most prolific inventors in history with 1,093 US patents in his name. He invented among others the phonograph, the automatic telegraph, the movie camera, the alkaline battery and the long-lasting, practical electric light bulb.
R1b-U106 branch (Germanic)
")
Larmuseau et al. (2013) tested the Y-DNA of three living members of the House of Bourbon, one descending from Louis XIII of France via King Louis Philippe I, and two from Louis XIV via Philip V of Spain. They concluded that all three men share the same STR haplotype and belonged to haplogroup R1b-U106 (Z381* subclade). These results contradict earlier studies by Lalueza-Fox et al. (2010), who had analysed the DNA from a handkerchief dipped in the presumptive blood of Louis XVI after his execution, as well as by Charlier et al. (2012), who tested the DNA of the severed head which allegedly belonged to Henry IV of France. Both of these studies had identified the remains as belonging to haplogroup G2a. All kings of France being descended in patrilineal line from Robert the Strong (820-866), unless a non-paternity event happened some time before Louis XIII, it can be assumed that all kings of France belonged to the same R1b-Z381 lineage. The House of Bourbon also includes all the kings of Spain from Philip V (1683-1746) to this day with King Juan Carlos, all the kings of the Two Sicilies, the grand dukes of Luxembourg since 1964, and of course all the dukes of Orléans and the dukes of Bourbon.

")
The lineage of the House of Wettin was identified as R1b-U106 > Z2265 > Z381 > Z156 > Z305 > Z307 > Z304 > DF98 > S18823 > S22069 > Y17440 > A6535 by the testing of two known descendants of the Wettin line by Brad Michael Little. The results confirmed that the two men, who share a common ancestor in Francis, Duke of Saxe-Coburg-Saalfeld, possess the same haplogroup and haplotype. Members of the House of Wettin include the Kings Edward VII, George V, Edward VIII and George VI of the United Kingdom, all the Kings of the Belgians, the Kings of Portugal from 1853 to 1910, the Kings of Bulgaria from 1887 to 1946, several Kings of Poland and Grand Dukes of Lithuania, the Margraves of Meissen from 1075 to 1423, the Electors of Saxony from 1423 to 1806, the Kings of Saxony from 1806 to 1918, and the rulers of the numerous smaller Saxon duchies.

")
Clan Gordon is a Scottish clan originating in the 12th century in the Scottish Borders region. The main branch descends from Elizabeth Gordon, daughter of Sir Adam Gordon, who married Alexander Seton in 1408. Their son was Alexander Gordon, 1st Earl of Huntly (d. 1470), whose patrilineal line was therefore Seton/Seaton and not the original Gordon lineage, was the progenitor of all the subsequent chiefs of clan. His descendants later became Marquesses of Huntly (1599-present), as well as Dukes of Gordon from 1684 to 1836. A cadet branch of the Huntly became Earls of Aberdeen (1682), then Marquesses of Aberdeen and Temair from 1916. The Gordon DNA Project identified the aristocratic Seton-Gordon line as R1b-U106 > Z2265 > S12025 > FGC52782 > FGC15048 (and matching people with the surname Seaton). The parent clade (FGC52782) has been found in Scandinavia and points to a common ancestor living a bit over 1000 years ago, suggesting that the Seton-Gordon line was of Viking origin.

The main branch of Scottish Clan Boyd, whose clan chief has held the title of Lord Boyd (1454) then of Earl of Kilmarnock from 1661 to 1746, belongs to R1b-U106 > Z381 > S1684 > U198 > S15627 > DF89 > FGC12770 > FT69836 > JFS0024 according to the Boyd DNA Project.

Benjamin Franklin (1706-1790), a polymath and one of the Founding Fathers of the United States, whose head has featured on the $100 bills since 1914, is a probable member of haplogroup R1b-U106 > Z18 > DF95 according to the testing of relatives. Ben Franklin was a political theorist, a statesnman, a diplomat, and a scientist, among others. He is also known for being the inventor of the lightning rod, bifocals, and the Franklin stove.

John Witherspoon (1723-1794) was one of the Founding Father of the United States and the president of Princeton University from 1768 to 1794. He was the only active clergyman and the only college president to sign the Declaration of Independence. Based on results from the Witherspoon DNA Project was a member of R1b-U106 > S263 > L48 > L47 > Z159 > Z4714 > BY35813 > BY56495.

John Rutledge (1739-1800) and his brother Edward Rutledge (1749-1800) were both Founding Fathers of the United States. Edward was the youngest signatory of the United States Declaration of Independence. John was one of the original Associate Justices of the Supreme Court and as its second Chief Justice. He also served as first governor of South Carolina after the Declaration of Independence, while he brother was the 10th governor. Descendant testing from the Rutledge/Routledge DNA Project determined that they belonged to R1b-U106 > S263 > L48 > L47 > L44 > L163 > Y23482 > Y23476 .

James K. Polk (1795-1849), the 11th president of the United States, was a member of L48 subclade based on the results from the Polk-Pollock DNA Project. He was a descendant of William Polk/Pollock (c. 1700-1757). Polk is chiefly known for extending the territory of the United States through the MexicanAmerican War; during his presidency, the United States expanded significantly with the annexation of the Republic of Texas, the Oregon Territory, and the Mexican Cession following the American victory in the MexicanAmerican War.

Franklin Pierce (1804-1869), the 14th president of the United States, was a member of R1b-U106 > Z381 > Z156 > S497 > DF96 based on the results from the Pierce DNA Project (North) (Group C). Pierce was a northern Democrat who believed that the abolitionist movement was a fundamental threat to the unity of the nation, he alienated anti-slavery groups by supporting and signing the KansasNebraska Act and enforcing the Fugitive Slave Act.

Nathan Bedford Forrest (1821-1877) was a prominent Confederate Army general during the American Civil War, renowned as a cavalry leader and military strategist. He was the only general on either side who began as a private. After the war he became the first Grand Wizard of the Ku Klux Klan, which has made him a controversial figure in American history. His lineage was identified (group B) in the Forrest/Forrester DNA Project, which makes him a member of R1b-U106 > S263 > S499 > L48 > Z9 > Z30 > Z349 > Z2 > Z7 > S5945 > FGC17344 > Y28576 > FGC51332.

According to the Grant DNA Project, Ulysses S. Grant (1822-1885), the 18th President of the United States and the military commander of the American Civil War, belonged to R1b-U106 > S263 > S499 > L48 > L47 > Z159 > S3249 > Y2924 > Y3264 > FGC8590 (descendant of Matthew Graunt).

William Howard Taft (1857-1930), the 27th President of the United States and the tenth Chief Justice of the United States, belonged to R1b-U106 > Z381 > Z301 >Z30 > Z338 > FGC1954 according to the results from the Taaffe DNA Project (descendant of Robert Taft, 1640-1725). Taft also served as 6th Solicitor General of the United States (1890-1892), Governor-General of the Philippines (1901-1903), 42th United States Secretary of War (1904-1908), and 1st Provisional Governor of Cuba (1906).

American journalist and writer Ernest Hemingway (1899-1961), who won the Nobel Prize of Literature in 1954, was a descendant of Ralph Hemingway who emigrated from England to Boston, MA. As such he must have belonged to R1b-U106 > Z381 > Z301 > L48 > L47 > Z159 > FGC15335 according to the Hemingway Y-DNA Project.

The two first human beings to have their whole genome sequenced, James D. Watson, the co-discoverers of the structure of DNA, and biologist Craig Venter both happen to be members of Y-DNA haplogroup R1b-S21 (U106), although they belong to distinct subclades (L48 > Z9 > Z30 for Watson, and L48 > L47 > L44 > L163 > L46 > L45 for Venter). Though both are American citizens, their patrilineal ancestors are respectively Scottish and German.
")
Actor Alec Baldwin (b. 1958) is a descendant of John Baldwin (1635-1683) of Stonington, Connecticut, which places him in the R1b-U106 cluster in the Baldwin Surname DNA Project. Nevgen assign his haplotype to R1b-U106 > Z381 > Z156 > BY12482 with 100% certainty. Baldwin won two Primetime Emmy Awards, three Golden Globe Awards, and seven Screen Actors Guild Awards, making him the male performer with the most SAG Awards in history.

The Academy Award nominated American actor and a playwright Woody Harrelson (b. 1961) belongs to haplogroup R1b-U106 > Z2265 > Z18 > Z372 > ZP91 > ZP158 > ZP161 > ZP262 according to the Harrelson DNA Project (descendant of Paul Harrelson from Denmark).
Other famous members of haplogroup R1b-U106
- Emir Abdelkader (1808-1883) : was an Algerian religious and military leader who led a struggle against the French colonial invasion in the mid-19th century. One of his descendants tested at the Syrian DNA project (kit BP18930) and was found to belong to R1b-U106 > Z381 > Z156 > S497 > DF96, which in Algeria could be of Vandalic (or possibly Gothic) origin.
- Charles Curtis (1860-1936) was an American attorney and Republican politician from Kansas who served as the 31st vice president of the United States from 1929 to 1933 under Herbert Hoover. He also previously served as the Senate Majority Leader from 1924 to 1929. Testing of relatives determined that he belonged to R1b-U106 > Z381 > Z301 > S1688 > U198 > S16994.
- Malcolm Gladwell (born 1963) : is an English-born Canadian journalist, author, and speaker. He has been a staff writer for The New Yorker since 1996. He has written five books, which were all on The New York Times Best Seller list. His haplogroup was revealved in Faces of America.
R1b-DF19 branch (Germanic)

Ringbauer et al. (2026) tested the genomes of Holy Roman Emperors Otto the Great and his great-nephew Henry II. Both were found to belong to R1b-P312 > DF19 > DF88 > Z17121 > FTA63331. By extension, other members of the Ottonian Dynasty also belonged to that haplogroup, including emperors Otto II and Otto III.
")
The Grant DNA Project identified the main lineage of Clan Grant descending from Duncan Grant of Freuchie (c. 1413-1485), 1st Laird of Freuchie and other early chiefs as belonging to R1b-P312 > DF19 > DF88 > Z17274. Later chiefs became Earls of Seafield (1701 to present), Barons Strathspey (1858 to present)
R1b-P312 branch (Celto-Germanic)
")
The ancestral lineage of Lowland Scottish Clan Armstrong appears to be R1b-P312 > FGC84729 > ZZ37 > ZZ38 > Z30597 > Z30600 > Z39305 > A9439 > FT275048 according to the Armstrong Y-DNA Project. The most famous member of the clan was probably the astronaut Neil Armstrong, who was the first person to walk on the Moon, and who took with him a piece of the Clan Armstrong tartan.

The influential Randolph family of Virginia belongs to R1b-P312 > Z39300 (aka PH2278) based on the results from the Randolph/Randall/Randle/Randol Surname Group (Albemarle County group). The family contributed numerous politicians, statesmen and military officers in the course of U.S. history. The most prominent carrier of the Randolph name was perhaps Peyton Randolph (1721-1775), who served as the the first President of the Continental Congress during the American Revolution. Edmund Randolph (1753-1813) was the seventh Governor of Virginia, the second Secretary of State, and the first United States Attorney General during George Washington's presidency. George W. Randolph (1818-1867) was a Confederate general who served for eight months in 1862 as the Confederate States Secretary of War during the American Civil War.

Woodrow Wilson, 28th President of the United States, belonged to haplogroup R1b-P312 > Z30597 based on the testing of a descendant of his grandfather, Judge James Wilson (1787-1850) at the Wilson DNA Project (Genetic Family ZZN). Wilson served as the president of Princeton University and as the 34th governor of New Jersey before winning the 1912 presidential election. As president, he oversaw the passage of progressive legislative policies unparalleled until the New Deal in 1933. He also led the United States into World War I in 1917, establishing an activist foreign policy known as Wilsonianism. He was the leading architect of the League of Nations.
R1b-L21 branch (Atlantic Celtic)

The O'Neill dynasty is a Gaelic Irish lineage that dominated the northern half of Ireland from the 6th to the 10th century. They claim descent from the 4th-century Irish king Niall of the Nine Hostages. According to the O'Neill DNA Project, the latter's probable lineage was R1b-L21 > DF13 > DF49 > Z2980 > Z2976 > DF23 > Z2961 > S645 > Z2965 > M222 > Y2605 > Y2841 > DF104 > DF109 (aka DF105), which split into 29 branches. Among these, it is believed that the S588 clade corresponds to the Cenél nEógain, the DF85 clade to the Cenél Conaill, and the A18726 clade to the Uí Briúin (royal dynasty of Connacht). The O'Neills have held the titles of Earl of Tyrone, Viscount O'Neill, Baron O'Neill, among others. Notable O'Neills include the American playwright and Nobel laureate Eugene O'Neill (1888-1953), the American businessman and founder of the O'Neill brand Jack O'Neill (1923-2017), the 4th Prime Minister of Northern Ireland Terence O'Neill (1914-1990), and the British economist Jim O'Neill, Baron O'Neill of Gatley (b. 1957).

The Dalcassians are a Gaelic Irish tribe who became powerful during the 10th and 11th centuries, when Brian Boru (941-1014) became High King of Ireland. He was the founder of the O'Brien dynasty, who were at various times in history Kings of Munster, Kings of Thomond (1118-1543), Earls of Thomond (1543-1726), Marquesses of Thomond (1726-1855), and Barons Inchiquin -1543-present), among others. According to the O'Brien DNA Project, most O'Brien members fall within the Dál gCais group belonging to R1b-L21 > DF13 > Z253 > Z2534 > L226, with a most recent common ancestor living about 1500 years ago.

Clan MacLean is one of the oldest clans in the Highlands, with origins going back at least to 852. The clan owned large tracts of land in Argyll as well as the Inner Hebrides. According to the MacLean DNA Project, the chiefly lines of Duart and Lochbuie belong to R1b-L21 > DF13 > DF1 > S5668 > FGC13499 > S5979 > S5982 > Z17816 > A1069 > Z17815, with a most recent common ancestor living about 1000 years ago.

Clan Gregor (or McGregor) is a Highland Scottish clan that claims an origin in the early 800s. The clan's most famous member is the folk hero Rob Roy MacGregor (1671-1734). The Clan is also known to have been among the first families of Scotland to begin playing the bagpipes in the early 17th century. The Clan Gregor DNA Project identified the main MacGregor line from Argyllshire as belonging to R1b-L21 > DF13 > Z39589 > L1335 > L1065 > Z16325 > S744 > S691 > S695 > BY144 > S690. The surname MacGregor was proscribed by Royal decree in Scotland from about 1650 to 1750 and many clan members changed their name to Murray. The clan chief has held the title of baronet Murray, then baronet MacGregor of MacGregor, since 1795. Scottish actor Ewan McGregor (b. 1971) could also belong to that haplogroup, although it hasn't been confirmed yet.

Clan Chattan is a confederaton of Highland Scottish clans. It was formed by descendants of the original clan, namely Clan Macpherson, Clan MacBean (or McBain) and Clan MacPhail, with the Clan Mackintosh and their cadet branches (Clan Shaw, Clan Farquharson, the Ritchies, and Clan MacThomas), and other clans unrelated by blood such as the MacGillivrays and the Davidson. According to the Clan Chattan DNA Project, most of the people with the original clan surnames fit within the R1b-L21 > DF13 > L1335 > S733 > Z16325 > S744 > Y16858 clade.

According to the Buchanan DNA Project, most members of the Highlands Scottish Clan Buchanan belong to haplogroup R1b-L21 > DF13 > L1335 > L1065 > S744 > S691 > S695. The clan is said to descend from Anselan O Kyan, a son of one of the kings of Ulster, who landed on the northern coast of Argyll, near Lennox, about the year 1016 and helped Malcolm II of Scotland in repelling the Danes. Some of the most notable clan members include the 15th President of the United States, James Buchanan (1791-1868), and American political commentator Pat Buchanan (b. 1938), who descends from the medieval clan chiefs.

Results from the Clan Cameron DNA Project indicate that the vast majority of participants of Highland Clan Cameron belong to R1b-L21 > DF13 > Y16233 > A2070 > Y16765 > A7298 > A6138. This includes several cadet branches. However the clan chiefs, Cameron of Lochiel, haven't been tested yet. The origins of the clan are unclear, but one hypothesis is that Donal Dubh, the first chief of Clan Cameron was descended either from the Macgillonies. Genetic genealogy confirms this link as members of Clan MacGillonie carry the same haplotype as the Cameron. British Prime Minister David Cameron and Canadian film director James Cameron probably belong to the same lineage, but it hasn't been confirmed yet.
")
Clan Campbell is historically one of the largest and most powerful of the Scottish Highland clans. The clan chief have held the title first of Lord Campbell (from 1445), then Earl of Argyll (1457), Marquess of Argyll (1641) and lastly Duke of Argyll from 1701 to this day. Another branch became the Earls of Loudoun (1633-1786). The Clan Campbell DNA Project identified the main Campbell line from Argyllshire as belonging to R1b-L21 > DF13 > Z39589 > L1335 > L1065 > FGC10125 > FGC10117, with a common patrilineal ancestor who lived about 1000 years ago.
")
Clan Drummond is a Highland Scottish clan whose clan chief were granted the title of Lord Drummond of Cargill (1488), then of Earl of Perth (1605 to this day). From 1716 to 1800, a Jacobite branch also held the title of Duke of Perth. The Drummond DNA Project determined that the Drummond Peerage belonged to R1b-L21 > DF13 > L513 > S5668 > A7 > Z17623 > S5982 > FGC30224 > ZS4584 > BY4017.

According to the Stewart Stuart DNA Project, the House of Stuart, who ruled Scotland from 1371, then also England and Ireland from 1603 until 1707, belongs to R1b-L21 > DF13 > Z39589 > DF41/S524 > Z43690 > S775 > L746 > S781. The most prominent members were King Robert II of Scotland, Kings James I, Charles I, Charles II and James II of England & Ireland. The Confederate States Army general J. E. B. Stuart (1833-1864) descends from the Baronet of Castle Stuart and therefore belonged to the same lineage. Formula One champion Jackie Stuart (b. 1939) desends from an illegitimate branch of King Robert III of Scotland, the Stewarts of Blackhall & Ardgowan.

James Scott, 1st Duke of Monmouth was the eldest illegitimate son of Charles II of England, Scotland, and Ireland with mistress Lucy Walter. He married Anne Scott, 1st Duchess of Buccleuch and daughter of the 2nd Earl of Buccleuch. The firstborn son descending from this couple have been carrying the title of Duke of Buccleuch and Earl of Dalkeith (among others), and been the Chief of Clan Scott ever since. In 2012, the 10th Duke of Buccleuch showed that he shared the same Y chromosome as a distant Stuart cousin, thus confirming that Charles II was indeed Monmouth's father (see House of Stuart above). Other ducal lineages descended from illegitimate sons of King Charles II include the Dukes of Cleveland (extinct in 1891), the Dukes of Grafton, the Dukes of St Albans, and the Dukes of Richmond (combined with the Dukedoms of Lennox and Gordon). The Dukes of Berwick, who have been part of the Spanish nobility since 1707, descend from James FitzJames, the illegitimate son of King James II, and therefore also carry the Stewart Y-chromosomal line. The current Duke of Berwick also hold 3 other dukedoms (including Duke of Alba) and 35 other titles, making him the most titled aristocrat in the world.
")
Clan Mackenzie is a Highland Scottish clan descending from Kenneth Mackenzie, 1st of Kintail (d. 1304). The Mackenzie have possessed the titles of Earl of Seaforth from 1623 to 1781, and of Earl of Cromartie (1703-1746 and 1861 to present). The clan's main branch, including confirmed descendants of Alexander Mackenzie of Kintail back in the 15th century, belong to R1b-L21 > DF13 > DF1 > S5668 > A7 > S7834 (aka S7828), with a common patrilineal ancestor who lived about 1200 years ago, based on the results from the Mackenzie DNA Project.
")
The lineage of the clan chiefs of Highland Scottish Clan MacLaren belong to to R1b-L21 > DF13 > Z39589 > L1335 > L1065 > S744 > S764 > Y16252 > BY23333 according to the Clan MacLaren DNA Project.

American statesman and lawyer and Founding Father Roger Sherman (1721-1793) probably belonged to R1b-L21 > DF13 > FGC5494 based on the results from the Sherman DNA Project. He is the only person to have signed all four great state papers of the United States: the Continental Association, the Declaration of Independence, the Articles of Confederation, and the Constitution. Union general William Tecumseh Sherman (1820-1891), who played a key role during the American Civil War, shared a common ancestor with Roger Sherman in the person of Henry Sherman (1511-1590) from Dedham, Essex. So both men would share the same paternal haplogroup. The M4 Sherman, the most widely used medium tank by the Allies in World War II, was named after General Sherman.

American Founding Father Patrick Henry (1736-1799) belonged to R1b-L21 > DF13 > DF1 > S6365 > BY16 > Z16372 according to relative testing at the Henry DNA Project (lineage R1b-06). Henry is best known for his declaration to the Second Virginia Convention (1775): "Give me liberty, or give me death!" He served as the first and sixth post-colonial Governor of Virginia.

Henry Clay (1777-1852) was an American attorney and statesman who represented Kentucky in both the Senate and House and served as 9th Secretary of State. He was part of the so-called Great Triumvirate, along with Daniel Webster and John C. Calhoun, who dominated American politics for much of the first half of the 19th century. Clay ran for U.S. president in 1824, 1832, and 1844 and received the electoral votes each time, but nevertheless lost the elections. Many descendants of his great-great-grand-father Charles Clay (d. 1686) from Virginia tested at the Clay DNA Project (lineage 01), which fits into haplogroup R1b-L21 > DF13 > FGC11134 > CTS4466 > A541 > A195 > BY55890.

The 12th president of the United States, Zachary Taylor (1784-1850) belonged to haplogroup R1b-L21 > DF13 > Z39589 > L1335 > L1065 > S764 > BY3148 > FGC18451 based on the results from the Taylor DNA Project (Group 06), and is ultimately descended from Sir John Taylor (1327-1377), a Knight to King Edward III. Taylor previously was a career officer in the United States Army, rose to the rank of major general and became a national hero as a result of his victories in the MexicanAmerican War. His son, Richard Taylor (1826-1879) was a Lieutenant General in the CSA and became army commander in the Trans-Mississippi Theater.

William Ewart Gladstone (1809-1898), a British Liberal politician who served a record four times as Prime Minister under Queen Victoria, belonged to R1b-L21 based on a comparison of his genealogy with the results from Gladstone DNA Project. His deep clade would be R1b-L21 > DF13 > FGC5494 > FGC5561 > Z16503 > Z16504 > Z16502 > Z17653 > L1444 > BY23824.

The analysis of his descendants's Y-chromosomal DNA confirmed that Joseph Smith (1805-1844), the founder of Mormonism and the Latter Day Saint movement, belonged to haplogroup R1b-M222 (R1b-L21 > DF13 > DF49 > Z2980 > Z2976 > DF23 > Z2961 > S645 > Z2965 > M222).

Rutherford B. Hayes (1822-1893), the 19th president of the United States, most probably belonged to haplogroup R1b-L21 > DF13 > L513 > S5668 > Z16357 > Z16854 > ZS349 based on the results from the Hayes DNA Project (Group H, descendant of George Hayes, 1655-1725). Hayes was a lawyer and staunch abolitionist. At the start of the Civil War, he joined the Union Army as an officer. He was wounded five times, earned a reputation for bravery in combat and was promoted to the rank of brevet major general.

George B. McClellan (1826-1885) was an American soldier, civil engineer, railroad executive, and politician who served as the 24th Governor of New Jersey. As major general during the American Civil War he played an important role in raising a well-trained and organized army. He was Commanding General of the Union Army for 5 months. His paternal lineage was identified through the McClellan Y-DNA Project as a descendant of Brig. General Samuel McClellan, making him a member of haplogroup R1b-L21 > DF13 > Z39589 > DF49 > DF23 > Z2961 > M222.

The American financier and banker J.-P .Morgan (1837-1913) descended from Sargent Miles Morgan who migrated from Wales to Massachusetts. This places him in Group 23 in the Morgan Surname Y-DNA Project corresponding to haplogroup R1b-L21 > DF13 > Z39589 > FGC43861 > BY2868 > BY132302.

William McKinley (1843-1901), the 25th president of the United States, probably belonged to haplogroup R1b-L21 > DF13 > L1335 > S764 based on the results from the MacKinlay DNA Project (descendant of James McKinley b. 1670 in Callender, Perthshire). During his presidency, McKinley led the nation to victory in the SpanishAmerican War, raised protective tariffs to promote American industry, and kept the nation on the gold standard in a rejection of the expansionary monetary policy of free silver.

Warren G. Harding (1865-1923), the 29th president of the United States, belonged to R1b-L21 > DF13 > Z39589 > DF49 > DF23 > ZP75 based on the results from the Harden/Harding DNA Project. He descended from Richard Harding (1583-1657) from Denbury, Devon, England and his son Stephen (1623-1698) who migrated to Braintree, Massachussets and died in Rhode Island.

The forensic analysis of the skeletal remains of Che Guevara (1928-1967), the Argentine Marxist revolutionary and major figure of the Cuban Revolution, revealed that he belonged to haplogroup R1b-L21.

One of the greatest boxers of all time, Muhammad Ali (1942-2016), born Cassius Marcellus Clay Jr, belonged to haplogroup R1b-L21 (deep clade DF13 > Z253 > Z2185 > Z2186 > L1066 > Z40246 > FGC53689). His Y-DNA haplogroup was determined by Big Y testing of his brother, Rahaman Ali.
")
Actor Matt LeBlanc (b. 1967), well known for playing the character of Joey Tribbiani in the TV series Friends and its sequel Joey, has a genealogy that can be traced back to Daniel Leblanc (1626-1695) who migrated from France to Port-Royal in Acadia (modern Nova Scotia). The same individual is listed in the Normandy Y-DNA Project as a member of R1b-L21 > DF63 > BY592. Geni.com mentions his haplogroup as R1b-M269.
Other famous members of R1b-L21
- Henry Louis Gates (subclade M222): an American literary critic, educator, scholar, writer, and editor. He was the first African American to receive the Andrew W. Mellon Foundation Fellowship. He has received numerous honorary degrees and awards for his teaching, research, and development of academic institutions to study black culture. He is the host of the PBS television series Faces of America and Finding Your Roots, which use traditional genealogical research and genetics to discover the family history of well-known Americans, including their haplogroups.
- Bill O'Reilly (subclade M222): an American television host, author, journalist, syndicated columnist, and a political commentator. He is the host of the political commentary program The O'Reilly Factor on the Fox News Channel. His Y-haplogroup was revealed by the PBS television series Finding Your Roots.
- Bill Maher (subclade M222): an American comedian, writer, producer, political commentator, actor, media critic, and television host. As a television host, he is well known for the HBO political talk show Real Time with Bill Maher. His Y-haplogroup was revealed by the PBS television series Finding Your Roots.
- Rory Bremner (subclade M222): a Scottish impressionist and comedian, noted for his work in political satire and impressions of British public figures.
- Adrian Grenier (subclade M222): an American actor, producer, director and musician. He is best known for playing Vincent Chase in the HBO series Entourage.
R1b-U152 branch (Italo-Celtic)

Based on the data from the Habsburg Family Project, there is a high likelihood that the haplogroup of the House of Habsburg was R1b-U152 > L2 > Z41150 > DF90 > FGC59564. The Habsburg originated in the eponymous village in northern Switzerland. They first became Kings of the Romans in 1273, obtained thenf Dukes of Austria from 1278, Kings of Spain from 1516, Kings of Hungary and Croatia from 1526... The Habsburgs provided all the Holy Roman Emperors but one from 1440 until the dissolution of the empire in 1806. However the senior Y-DNA line of the Habsburgs ended with Emperor Charles VI (1685-1740), father of Empress Maria Theresa. Check also Haplogroups of European kings and queens for mtDNA lines of many Habsburg family members.

While testing for paternal relatives of Richard III of England to confirm the identity of his presumed remains (see Haplogroup G2a), researchers found that three modern relatives with the surname Somerset and descended from House of Lancaster all belonged to haplogroup R1b-U152 (x L2, Z36, Z56, M160, M126 and Z192). Although this points to a non-paternity at some time in the Plantagenet lineage, it is likely that most if not all Dukes of Beaufort, and possibly most Plantagenets monarchs outside the House of York belonged to R1b-U152.
")
Clan Erskine, a Lowland Scottish clan from Renfrewshire, apparently belongs to R1b-U152 > Z36 > BY1328 > BY2151 > A7992 > A8001 > BY32396 based on at least one descendant from John Erskine, 19th Earl of Mar (15581634) at the Erskine/Askey Surname Project. The Erskines have held the title of Earl of Mar since 1404, of Earl of Kellie since 1619, and of Earl of Buchan since 1640.

FamilyTreeDNA collaborated with the Armed Forces DNA Identification Laboratory The first President of the United States to identify the paternal line of the First President of the United States, George Washington (1732-1799). The study determined that George Washington belonged to R1b-U152 > Z36 > Y17161 > BY1328 > BY2151 > A7992.

Abraham Lincoln (1809-1865), the 16th president of the United States, appears to have belonged to R1b-U152 > L2 > Z142 > Z150 > S20376 > S20376. The Lincoln DNA Project tested several descendants from Samuel "the weaver" Lincoln, who was Abraham Lincoln's great-great-great-great-grandfather, and they all share the same haplotype.

Grover Cleveland (1837-1908), the 22nd and 24th president of the United States, could have belonged to R1b-L20, a subclade of L2, based on the numerous results from the Cleveland DNA Project.
")
The American singer, songwriter and dancer Michael Jackson (1958-2009), who was known as the "King of Pop", and he is regarded as one of the most significant cultural figures of the 20th century, belonged to haplogroup R1b-U152 > L2 > Z258 > Z367 > L20 > BY203086 > FT121435 > FT121274. His Y-DNA was determined by Big Y testing of a relative descending from Michael's paternal great-grandfather Israel Nero Jackson Sr (1838 - 1934) at Family Tree DNA. Not much is known about this deep clade except that it is also found in northern France.

Actor Kevin Costner (b. 1955) descends from George Adam Costner (Kastner) from Edenkoben, Rhineland-Palatinate, Germany, and his father Andreas Kastner. This places him in Group 2 in the Kastner Y-DNA Project, who belong to R1b-U152 > L2 > A19725 > BY39321.
")
American-Canadian actor Matthew Perry (b. 1969), most famous for playing Chandler Bing in the sitcom Friends, probably belongs to haplogroup R1b-U152 > L2 > Z49 > Z142 > CTS2819 based on results from members of the Perry Y-DNA Project (Family 7). He is a descendant of John Perry (1604-1674) and his father John Perry (1588-1621) from Hampshire, England.
Other famous members of R1b-U152
- Jörg Jenatsch (1596-1639) : was a Swiss political leader during the Thirty Years' War, one of the most striking figures in the troubled history of the Grisons in the 17th century.
- Mario Batali : is an American chef, writer, restaurateur and media personality. He is an expert on the history and culture of Italian cuisine, including regional and local variations. His haplogroup was revealved in Faces of America.
R1b-DF27 branch (Celtic)
")
Clan Bruce is a Lowlands Scottish clan, which was a Royal House in the 14th century, producing two kings of Scotland (Robert the Bruce and David II of Scotland), and a disputed High King of Ireland, Edward Bruce. The Bruces were first Lords of Annandale from 1124 until King Robert the Bruce conferred the title to his nephew, Thomas Randolph. Robert the Bruce successfully fought off the English in the First War of Scottish Independence and is now revered in Scotland as a national hero. Other clan members later became Barons of Clackmannan, Lords Bruce of Kinloss (1608), Earls of Elgin (1633) and Earls of Kincardine (1647), titles that they keep to this day. The Bruce DNA Project determined that the noble lineage of Clan Bruce belongs to R1b-DF27 > ZZ12 > Z46512 > FGC78762 > ZZ41 > S7432.
")
Highland Scottish Clan Murray and Clan Sutherland both descend from a Flemish nobleman by the name of Freskin, who settled in Scotland during the reign of King David I and was granted lands in West Lothian and the ancient Pictish kingdom of Moray (which would become known as Sutherland). Freskin's descendants were designated by the surname de Moravia ("of Moray" in the Norman language), which later became 'Murray'. Freskin's great-grand-son was William de Moravia (c. 12101248), became 1st Earl of Sutherland, a title that the clan chief who keep until 1535, when it passed to Clan Gordon. Clan Murray descends from William's cousin. They would acquire the title of Earl of Tullibardine (1606), Earl of Atholl (1629), Marquess of Atholl (1676), and ultimately Duke of Atholl (1703) to this day. The second son of the 1st Marquess of Atholl and his descendants would become Earls of Dunmore. According to the Sutherland DNA Project, the ancestral Sutherland line belongs to R1b-DF27 > ZZ12 > FGC23071 > FGC23066 > BY48361 > BY130907 > BY67446 and has Y-chromosomal matches in modern Flanders, confirming Freskin's origins. It is believed that Clan Douglas also descends from Freskin and the biggest cluster in the Douglas DNA Project indeed matches the Sutherland and Murray haplotype.
")
According to the Boyle DNA Project, the Earls of Glasgow belongs to R1b-DF27 > Z196 > SRY2627 > Z206 > BY3222. The Earl is the hereditary clan chief of Clan Boyle, whose ancestors came from the Norman town of Beauville near Caen.

John Hancock (1737-1793) was an American merchant, statesman, prominent Patriot of the American Revolution and signatory of the Declaration of Independence. He served as president of the Second Continental Congress and was the first and third Governor of Massachusetts. He was a descendant of
John Hancock (1506-1570) from Chesterfield, Derbyshire. Another branch descending from that ancestor tested on the Hancock DNA Project (Group 01) and was found to belong to R1b-DF27 > Z195 > Z198 > Z292 > Z262 > Z201 > Z202 > Z205 > Z206 > Z208. Another notable descendant of the same John Hancock of Chesterfield was Union general Winfield Scott Hancock (1824-1886). Known to his Army colleagues as "Hancock the Superb", he was noted in particular for his personal leadership at the Battle of Gettysburg in 1863.

The House of Bernadotte, which has been the royal house of Sweden since in 1818, and also of Norway between 1818 and 1905 belongs to R1b-DF27 > Z195 > Z272 > S450 > Z295 > S25783 > CTS4065 > S16864 > A5000 > FGC39239 according to the testing of a cousin of King Gustaf V. The dynasty was founded by Jean Bernadotte, a former marshal of Napoleon, who was elected the heir-presumptive to the childless King Charles XIII of Sweden, and became King Charles XIV John. Bernadotte was a native of Pau in Gascony, where this haplogroup is indeed common today.

Commodore Matthew Calbraith Perry (1794-1858), the man who forced Japan to open its ports to Western ships, which eventually caused the downfall of the Tokugawa shogunate and the Meiji restoration, probably belonged to haplogroup R1b-DF27 > Z196 > Z209 > Z295 based on shared genealogy (descendant of Edward Perry 1630-1695 from Devon, England) with members of the Perry Y-DNA Project (Family 11).

Braxton Bragg (1817-1876) was an American army officer during the Second Seminole War and MexicanAmerican War and later a Confederate army officer who served as a general in the Confederate Army during the American Civil War. His most important role was as commander of the Army of Mississippi. His fatherline was identified in the Bragg DNA Project (Family 2, descending from Joseph B. Bragg), making of him a member of R1b-DF27 > Z196 > SRY2627 > Z208.
")
The Austrian composer, music theorist and painter Arnold Schönberg (1874-1951) belonged to Y-haplogroup R1b according to his profile on Geni. According to the Schönberg DNA Project his lineage (grandson of Abraham Schoenberg, born in 1812 in Szécsény) falls under the BY16148 clade downstream of DF27.

George H.W. Bush (1924-2018), the 41st president of the United States, and his son George W. Bush (b. 1946), the 43rd president, apparently belong to haplogroup R1b-DF27 > Z196 > Z209 > CTS4065 > S16864 based on the results from the Bush DNA Project (group A). They descend from Reynold Bush (1600-1686) who emigrated from England (Fering Parish, Essex) to the Massachusetts colony about 1640.
Undetermined R1b branch
")
Yehia Z Gad and his team at the Ancient DNA lab of the National Museum of Egyptian Civilization in Cairo retrieved the DNA of several members of the 18th Dynasty of Egypt (c. 15501295 BCE), which included Amenhotep I to III, Thutmose I to IV, Hatshepsut, Akhenaten and Tutankhamun. The Y-DNA analysis established that The royal male lineage belonged to Y-haplogroup R1b.

Professor Tomasz Kozłowski tested the Y-DNA of Prince Janusz III of Masovia, Duke of Czersk, Warsaw, Liw, Zakroczym and Nur. He was a direct descendent of the Piast Dynasty, the first historical dynasty ruling over Poland, starting with Prince Mieszko I (c. 930992) and ending in 1370 with the death of King Casimir III the Great. The Piast family continued to rule over the Duchy of Masovia until 1526 and the Duchy of Silesia until 1675. Prof. Kozłowski announced that the prince belonged to haplogroup R1b, and therefore probably also other members of that royal lineage, including all the Dukes of Masovia (1138-1526), as long as no non-paternity event took place.

Bogdanowicza et al. (2009) tested the Y-chromosomal DNA and mitochondrial DNA of the exhumed remains of the Renaissance astronomer Nicolaus Copernicus. They established that he belonged to Y-haplogroup R1b-P310 and mt-haplogroup H.

Rogaev et al. (2009) tested the DNA of the presumed grave of Tsar Nicholas II of Russia and all his five children, and compared them against archival blood specimens from Nicholas II as well as against samples from descendants of both paternal and maternal lineages. The results unequivocally confirmed that the grave was the one of the last Russian Royal family. Nicholas II belonged to Y-haplogroup R1b and mt-haplogroup T2. Consequently, all Russian emperors of the Romanov dynasty since Peter III (1728-1762) also belonged to haplogroup R1b. This paternal lineage ultimately descends from the House of Oldenburg, which includes all the Kings of Denmark since Christian I (reigned from 1448) as well as several Kings of Norway, Sweden and Greece, and the current heirs to the British throne (Prince Charles and his son Prince William).

The great English naturalist Charles Darwin (1809-1882), who proposed the scientific theory of evolution and the process of natural selection, was a member of haplogroup R1b according to the test results from his great-great-grandson.

Kevin Bacon (b. 1958), an American actor and musician whose films include musical-drama film Footloose (1984), the controversial historical conspiracy legal thriller JFK (1991), the legal drama A Few Good Men (1992), the historical docudrama Apollo 13 (1995), and the mystery drama Mystic River (2003). Bacon has won a Golden Globe Award and three Screen Actors Guild Awards, and was nominated for a Primetime Emmy Award. The Guardian named him one of the best actors never to have received an Academy Award nomination.

The American actor Robert John Downey Jr. (b. 1965) had his Y-haplogroup was revealed by the PBS television series Finding Your Roots. His father adopted the surname Downey, but before that his paternal ancestors were called Elias, or Eliasovitz before they emigrated to the United States. He was nominated for the Academy Award for Best Actor and won him the BAFTA Award for Best Actor in a Leading Role in the movie Chaplin (1992), and won a Golden Globe Award for his role in the TV series Ally McBeal. He is also known for portraying the role of Marvel Comics superhero Iron Man.
Other famous members of haplogroup R1b
- Harry Connick Jr. (born 1967) : is an American singer, musician, and actor. He has sold over 28 million albums worldwide. Connick is ranked among the top 60 best-selling male artists in the United States by the Recording Industry Association of America.
- Charlie Rose (born 1942) : is an American television talk show host and journalist. Since 1991, he has hosted Charlie Rose, an interview show distributed nationally by PBS since 1993.
Read this article in other languages
Follow-up
Ask your questions and discuss about haplogroups on the Forum



 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")
 - Eupedia")






 in Europe")


 in Europe")
 in Europe")
 in Europe")



")

")

")
")












")


")

")




")





")
")


")
")













")


")



")

")
")
")
")




")

")
")






